

Adult hippocampal neurogenesis and cognitive flexibility - linking memory and mood
- PMID: 28469276
- PMCID: PMC6261347
- DOI: 10.1038/nrn.2017.45
Adult hippocampal neurogenesis and cognitive flexibility - linking memory and mood
Abstract
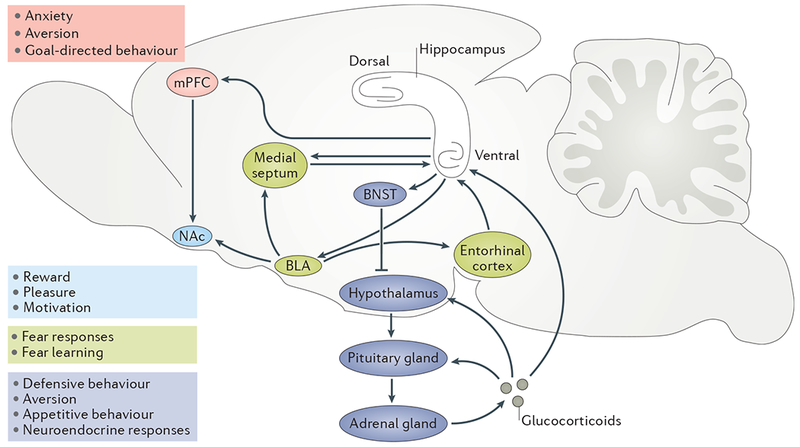
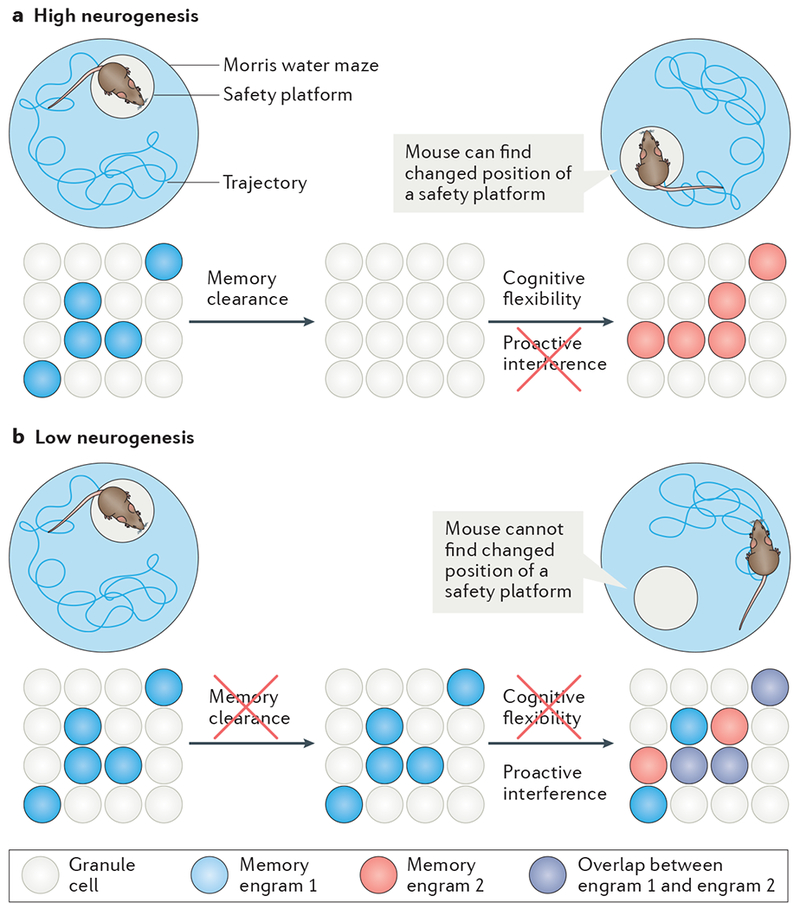
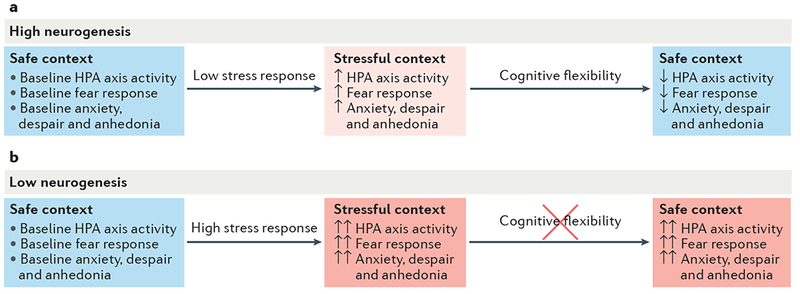
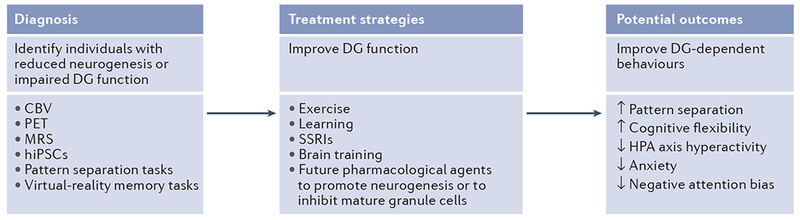
Adult hippocampal neurogenesis has been implicated in cognitive processes, such as pattern separation, and in the behavioural effects of stress and antidepressants. Young adult-born neurons have been shown to inhibit the overall activity of the dentate gyrus by recruiting local interneurons, which may result in sparse contextual representations and improved pattern separation. We propose that neurogenesis-mediated inhibition also reduces memory interference and enables reversal learning both in neutral situations and in emotionally charged ones. Such improved cognitive flexibility may in turn help to decrease anxiety-like and depressive-like behaviour.
Conflict of interest statement
Competing interests statement
The authors declare no competing interests.
Figures
References
-
- Taupin P & Gage FH Adult neurogenesis and neural stem cells of the central nervous system in mammals. J. Neurosci. Res. 69, 745–749 (2002). - PubMed
-
- Kjelstrup KB et al. Finite scale of spatial representation in the hippocampus. Science 321, 140–143 (2008). - PubMed
-
- Thompson CL et al. Genomic anatomy of the hippocampus. Neuron 60, 1010–1021 (2008). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
