

Hox11 Function Is Required for Region-Specific Fracture Repair
- PMID: 28470721
- PMCID: PMC5550340
- DOI: 10.1002/jbmr.3166
Hox11 Function Is Required for Region-Specific Fracture Repair
Abstract
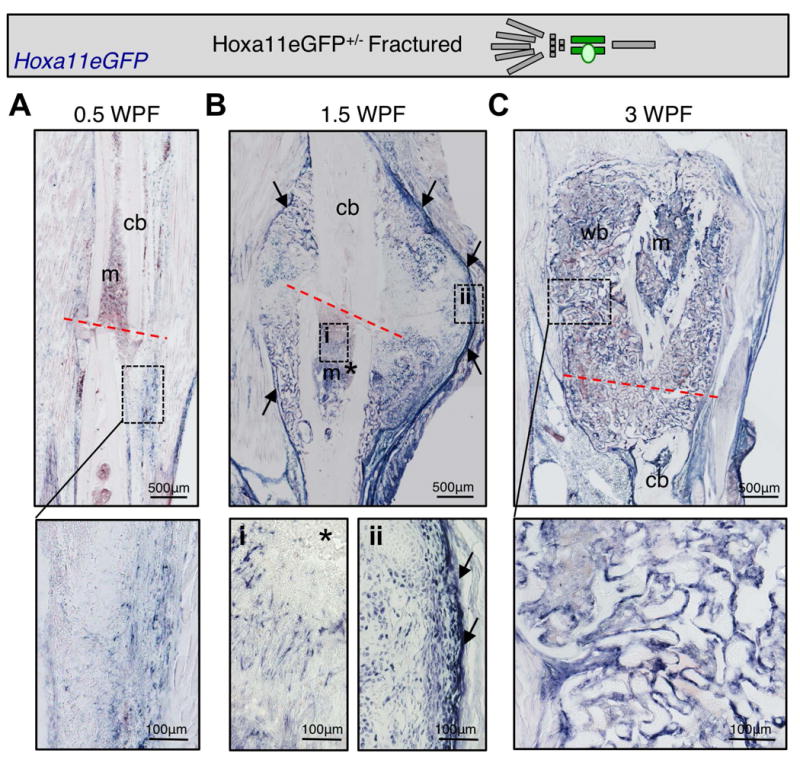
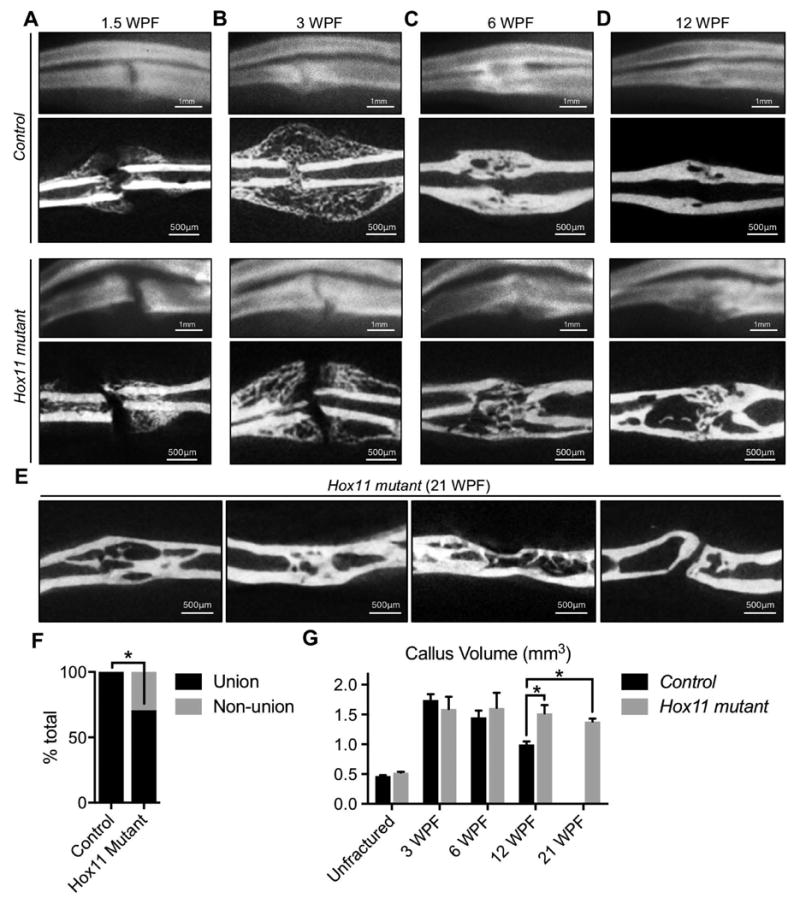
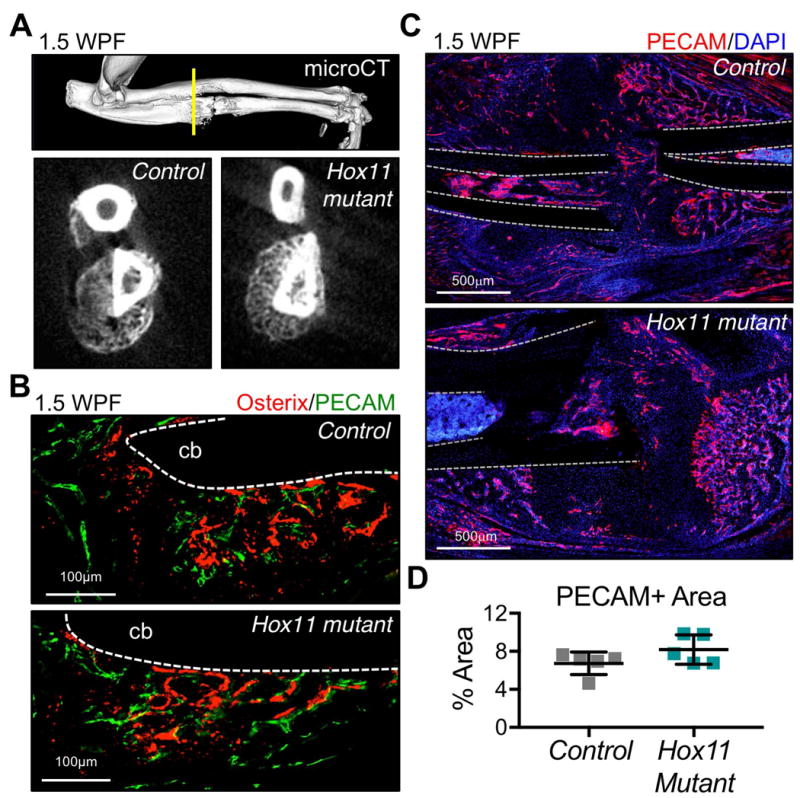
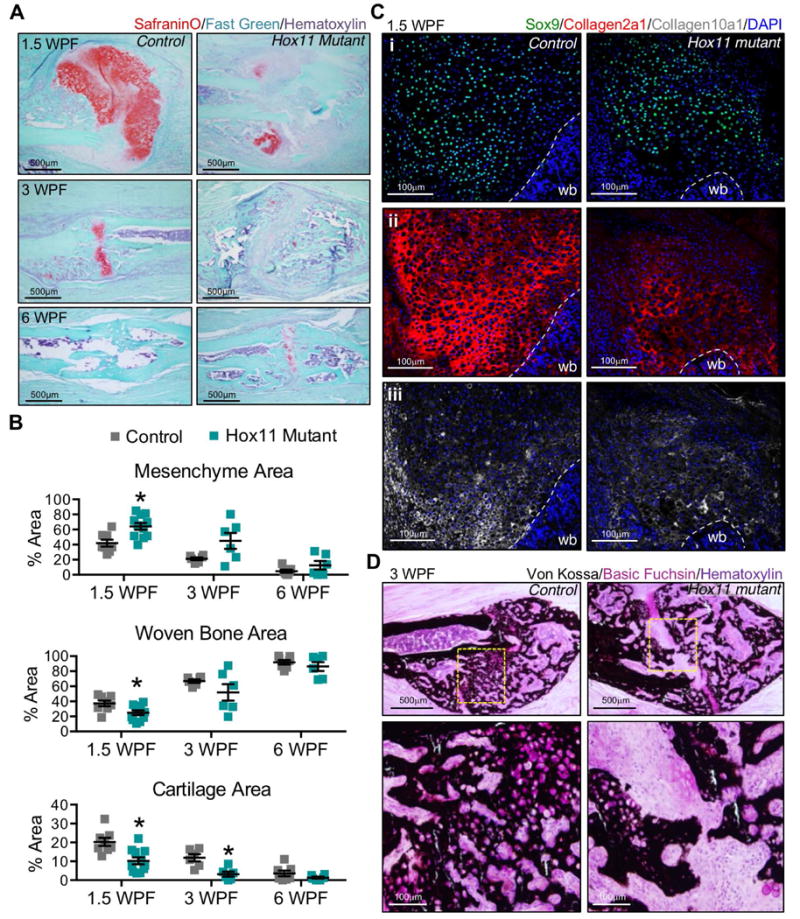
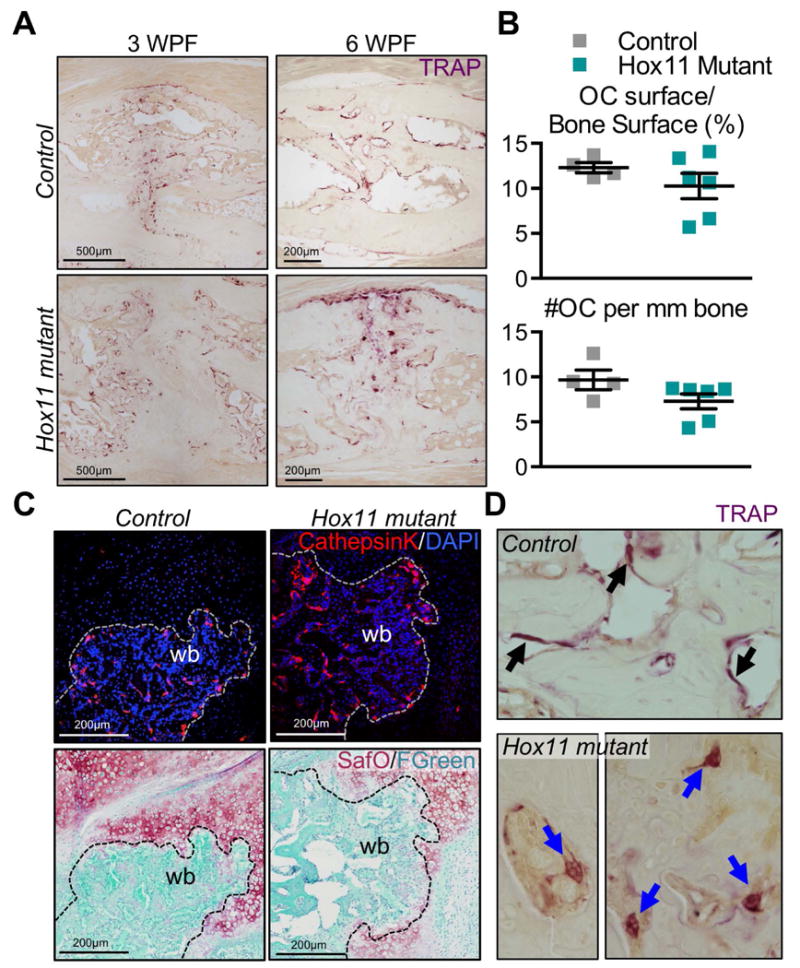
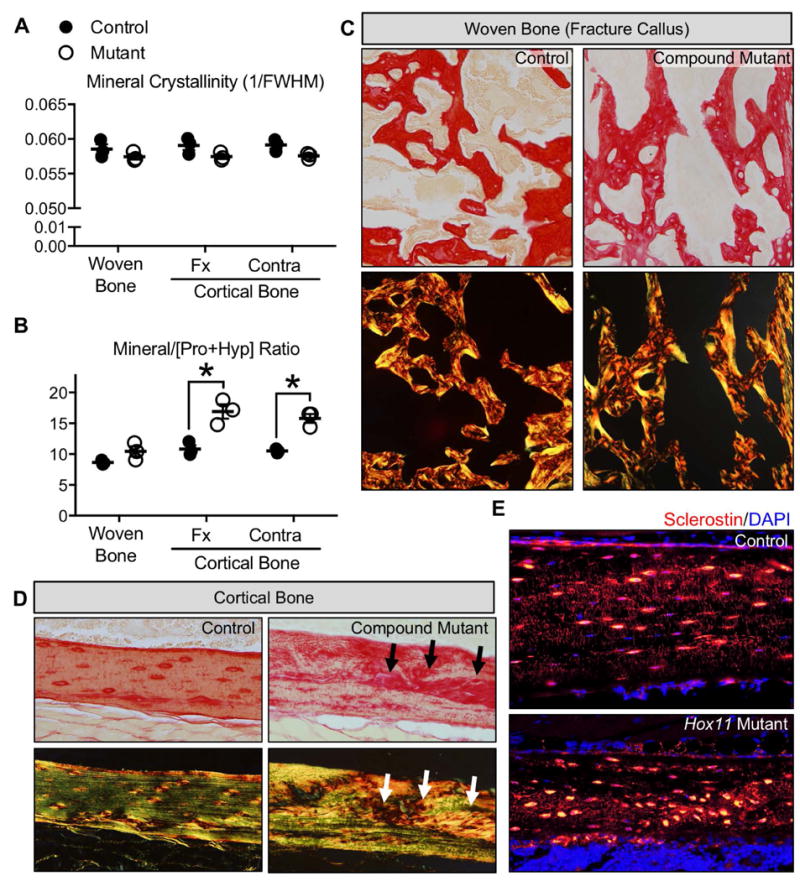
The processes that govern fracture repair rely on many mechanisms that recapitulate embryonic skeletal development. Hox genes are transcription factors that perform critical patterning functions in regional domains along the axial and limb skeleton during development. Much less is known about roles for these genes in the adult skeleton. We recently reported that Hox11 genes, which function in zeugopod development (radius/ulna and tibia/fibula), are also expressed in the adult zeugopod skeleton exclusively in PDGFRα+/CD51+/LepR+ mesenchymal stem/stromal cells (MSCs). In this study, we use a Hoxa11eGFP reporter allele and loss-of-function Hox11 alleles, and we show that Hox11 expression expands after zeugopod fracture injury, and that loss of Hox11 function results in defects in endochondral ossification and in the bone remodeling phase of repair. In Hox11 compound mutant fractures, early chondrocytes are specified but show defects in differentiation, leading to an overall deficit in the cartilage production. In the later stages of the repair process, the hard callus remains incompletely remodeled in mutants due, at least in part, to abnormal bone matrix organization. Overall, our data supports multiple roles for Hox11 genes following fracture injury in the adult skeleton. © 2017 American Society for Bone and Mineral Research.
Keywords: ENDOCHONDRAL OSSIFICATION-CARTILAGE; HOX GENES; MESENCHYMAL STROMAL/STEM CELLS; MOLECULAR PATHWAYS-DEVELOPMENT; SKELETAL INJURY/FRACTURE HEALING.
© 2017 American Society for Bone and Mineral Research.
Figures
Similar articles
-
Regionally Restricted Hox Function in Adult Bone Marrow Multipotent Mesenchymal Stem/Stromal Cells.Dev Cell. 2016 Dec 19;39(6):653-666. doi: 10.1016/j.devcel.2016.11.008. Epub 2016 Dec 8. Dev Cell. 2016. PMID: 27939685 Free PMC article.
-
Hox11 genes are required for regional patterning and integration of muscle, tendon and bone.Development. 2013 Nov;140(22):4574-82. doi: 10.1242/dev.096693. Epub 2013 Oct 23. Development. 2013. PMID: 24154528 Free PMC article.
-
Hox genes maintain critical roles in the adult skeleton.Proc Natl Acad Sci U S A. 2020 Mar 31;117(13):7296-7304. doi: 10.1073/pnas.1920860117. Epub 2020 Mar 13. Proc Natl Acad Sci U S A. 2020. PMID: 32170021 Free PMC article.
-
Fracture Healing in the Setting of Endocrine Diseases, Aging, and Cellular Senescence.Endocr Rev. 2022 Nov 25;43(6):984-1002. doi: 10.1210/endrev/bnac008. Endocr Rev. 2022. PMID: 35182420 Free PMC article. Review.
-
Site-Specific Fracture Healing: Comparison between Diaphysis and Metaphysis in the Mouse Long Bone.Int J Mol Sci. 2021 Aug 27;22(17):9299. doi: 10.3390/ijms22179299. Int J Mol Sci. 2021. PMID: 34502206 Free PMC article. Review.
Cited by
-
Widespread diversity in the transcriptomes of functionally divergent limb tendons.J Physiol. 2020 Apr;598(8):1537-1550. doi: 10.1113/JP279646. Epub 2020 Mar 30. J Physiol. 2020. PMID: 32083717 Free PMC article.
-
Generation of Hoxa11-3XFLAG and Hoxd11-3XFLAG alleles to investigate Hox11 genome-wide binding.Dev Biol. 2025 Aug;524:210-218. doi: 10.1016/j.ydbio.2025.05.013. Epub 2025 May 17. Dev Biol. 2025. PMID: 40389054
-
Niches for Skeletal Stem Cells of Mesenchymal Origin.Front Cell Dev Biol. 2020 Jul 10;8:592. doi: 10.3389/fcell.2020.00592. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32754592 Free PMC article. Review.
-
Hox5 genes direct elastin network formation during alveologenesis by regulating myofibroblast adhesion.Proc Natl Acad Sci U S A. 2018 Nov 6;115(45):E10605-E10614. doi: 10.1073/pnas.1807067115. Epub 2018 Oct 22. Proc Natl Acad Sci U S A. 2018. PMID: 30348760 Free PMC article.
-
Bone quality assessment of osteogenic cell cultures by Raman microscopy.J Raman Spectrosc. 2019 Mar;50(3):360-370. doi: 10.1002/jrs.5521. Epub 2018 Nov 28. J Raman Spectrosc. 2019. PMID: 37035410 Free PMC article.
References
-
- Bolander ME. Regulation of fracture repair by growth factors. Proceedings of the Society for Experimental Biology and Medicine Society for Experimental Biology and Medicine. 1992;200(2):165–170. - PubMed
-
- Einhorn TA. The cell and molecular biology of fracture healing. Clinical orthopaedics and related research. 1998;355(Suppl):S7–21. - PubMed
-
- Ferguson C, Alpern E, Miclau T, Helms JA. Does adult fracture repair recapitulate embryonic skeletal formation? Mech Dev. 1999;87(1-2):57–66. - PubMed
-
- Gerstenfeld LC, Cullinane DM, Barnes GL, Graves DT, Einhorn TA. Fracture healing as a post-natal developmental process: molecular, spatial, and temporal aspects of its regulation. Journal of cellular biochemistry. 2003;88(5):873–884. - PubMed
-
- Vortkamp A, Pathi S, Peretti GM, Caruso EM, Zaleske DJ, Tabin CJ. Recapitulation of signals regulating embryonic bone formation during postnatal growth and in fracture repair. Mech Dev. 1998;71(1-2):65–76. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
