

Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts
- PMID: 28472298
- PMCID: PMC5850832
- DOI: 10.1093/humrep/dex087
Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts
Abstract
Study question: How does a maternal diabetic hyperadiponectineamia affect signal transduction and lipid metabolism in rabbit preimplantation blastocysts?
Summary answer: In a diabetic pregnancy increased levels of adiponectin led to a switch in embryonic metabolism towards a fatty acid-dependent energy metabolism, mainly affecting genes that are responsible for fatty acid uptake and turnover.
What is known already: Although studies in cell culture experiments have shown that adiponectin is able to regulate lipid metabolism via 5'-AMP-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor α (PPARα), data on the effects of adiponectin on embryonic lipid metabolism are not available. In a diabetic pregnancy in rabbits, maternal adiponectin levels are elevated fourfold and are accompanied by an increase in intracellular lipid droplets in blastocysts, implying consequences for the embryonic hormonal and metabolic environment.
Study design, size, duration: Rabbit blastocysts were cultured in vitro with adiponectin (1 μg/ml) and with the specific AMPK-inhibitor Compound C for 15 min, 1 h and 4 h (N ≥ 3 independent experiments: for RNA analysis, n ≥ 4 blastocysts per treatment group; for protein analysis three blastocysts pooled per sample and three samples used per experiment). Adiponectin signalling was verified in blastocysts grown in vivo from diabetic rabbits with a hyperadiponectinaemia (N ≥ 3 independent experiments, n ≥ 4 samples per treatment group, eight blastocysts pooled per sample).
Participants/materials, setting, methods: In these blastocysts, expression of molecules involved in adiponectin signalling [adaptor protein 1 (APPL1), AMPK, acetyl-CoA carboxylase (ACC), p38 mitogen-activated protein kinases (p38 MAPK)], lipid metabolism [PPARα, cluster of differentiation 36 (CD36), fatty acid transport protein 4 (FATP4), fatty acid binding protein (FABP4), carnitine palmityl transferase 1 (CPT1), hormone-senstive lipase (HSL), lipoprotein lipase (LPL)] and members of the insulin/insulin-like growth factor (IGF)-system [IGF1, IGF2, insulin receptor (InsR), IGF1 receptor (IGF1R)] were analyzed by quantitative RT-PCR and western blot. Analyses were performed in both models, i.e. adiponectin stimulated blastocysts (in vitro) and in blastocysts grown in vivo under increased adiponectin levels caused by a maternal diabetes mellitus.
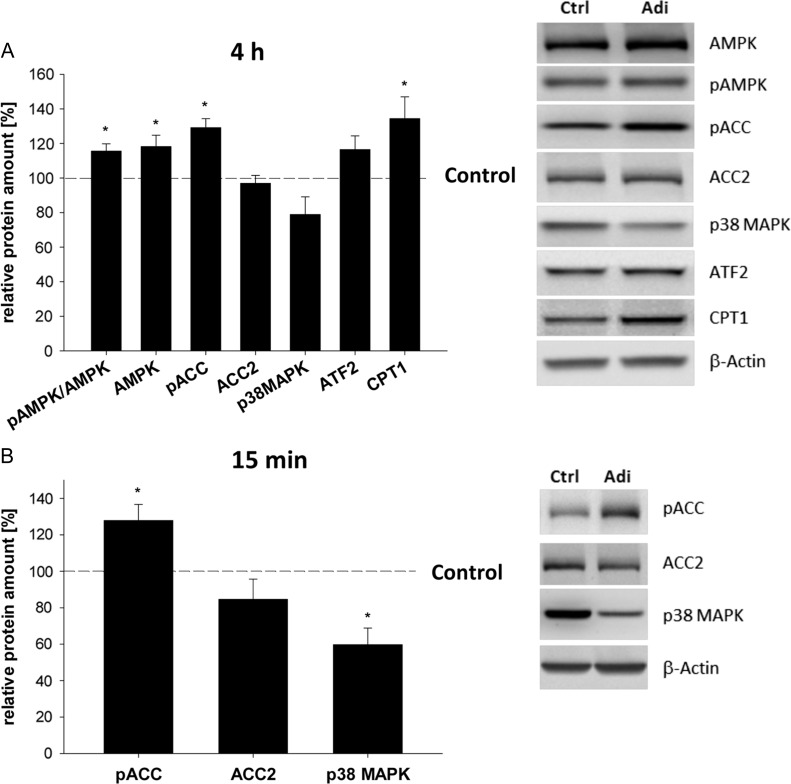
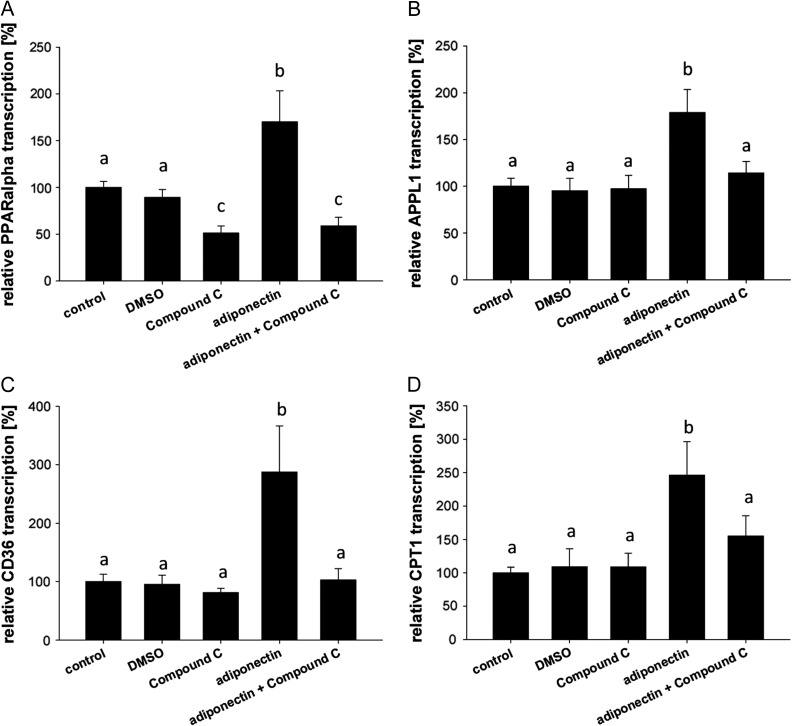
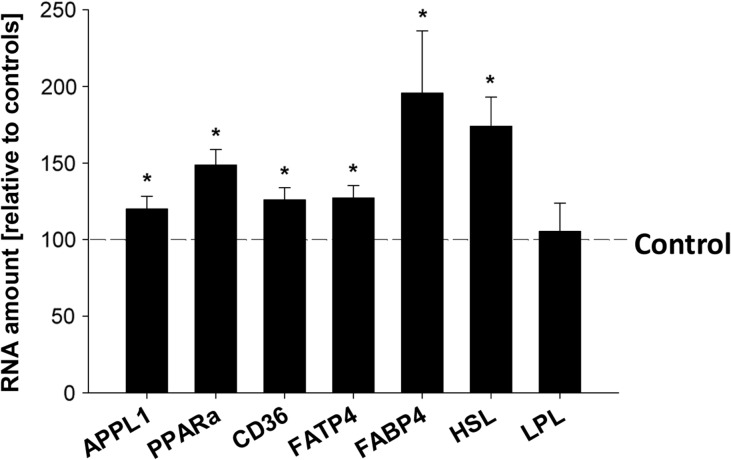
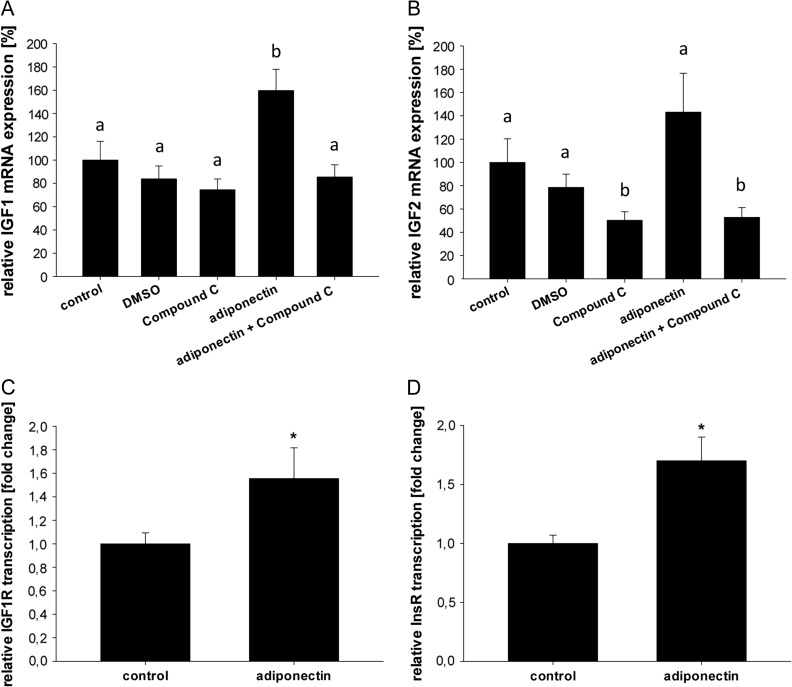
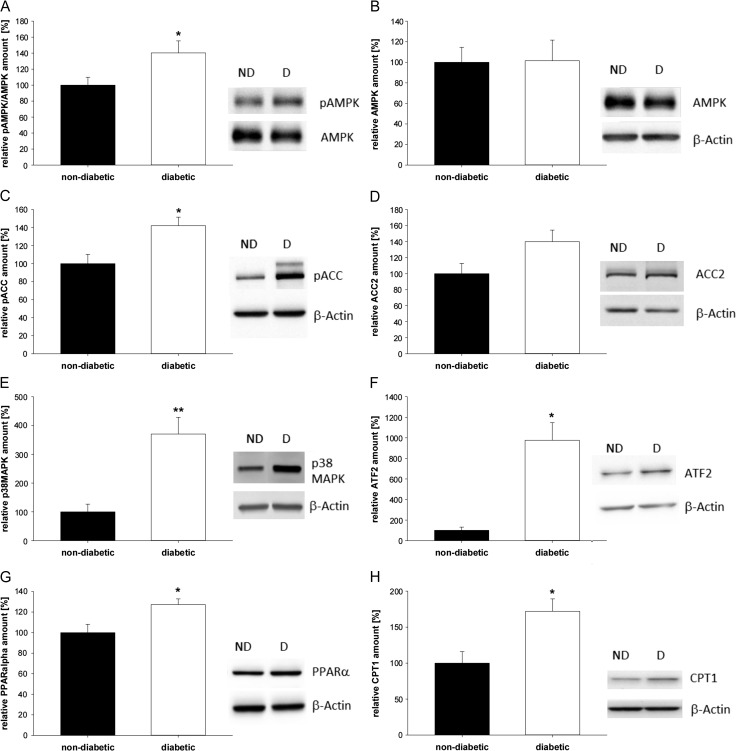
Main results and the role of chance: In both in vitro and in vivo models adiponectin increased AMPK and ACC phosphorylation, followed by an activation of the transcription factor PPARα, and CPT1, the key enzyme of β-oxidation (all P < 0.05 versus control). Moreover, mRNA levels of the fatty acid transporters CD36, FATP4 and FABP4, and HSL were upregulated by adiponectin/AMPK signalling (all P < 0.05 versus control). Under diabetic developmental conditions the amount of p38 MAPK was upregulated (P < 0.01 versus non-diabetic), which was not observed in blastocysts cultured in vitro with adiponectin, indicating that the elevated p38 MAPK was not related to adiponectin. However, a second effect of adiponectin has to be noted: its intensification of insulin sensitivity, by regulating IGF availability and InsR/IGF1R expression.
Large scale data: Not applicable.
Limitations reasons for caution: There are two main limitations for our study. First, human and rabbit embryogenesis can only be compared during blastocyst development. Therefore, the inferences from our findings are limited to the embryonic stages investigated here. Second, the increased adiponectin levels and lack of maternal insulin is only typical for a diabetes mellitus type one model.
Wider implications of the findings: This is the first mechanistic study demonstrating a direct influence of adiponectin on lipid metabolism in preimplantation embryos. The numbers of young women with a diabetes mellitus type one are increasing steadily. We have shown that preimplantation embryos are able to adapt to changes in the uterine milieu, which is mediated by the adiponectin/AMPK signalling. A tightly hormonal control during pregnancy is essential for survival and proper development. In this control process, adiponectin plays a more important role than known so far.
Study funding/competing interest(s): This work was supported by the German Research Council (DFG RTG ProMoAge 2155), the EU (FP7 Epihealth No. 278418, FP7-EpiHealthNet N°317146), COST Action EpiConcept FA 1201 and SALAAM BM 1308. The authors have no conflict(s) of interest to disclose.
Keywords: 5′-AMP-activated protein kinase; adiponectin; carnitine palmityl transferase 1; fatty acid uptake; lipid metabolism; peroxisome proliferator-activated receptor α; preimplantation embryo.
© The Author 2017. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology.
Figures
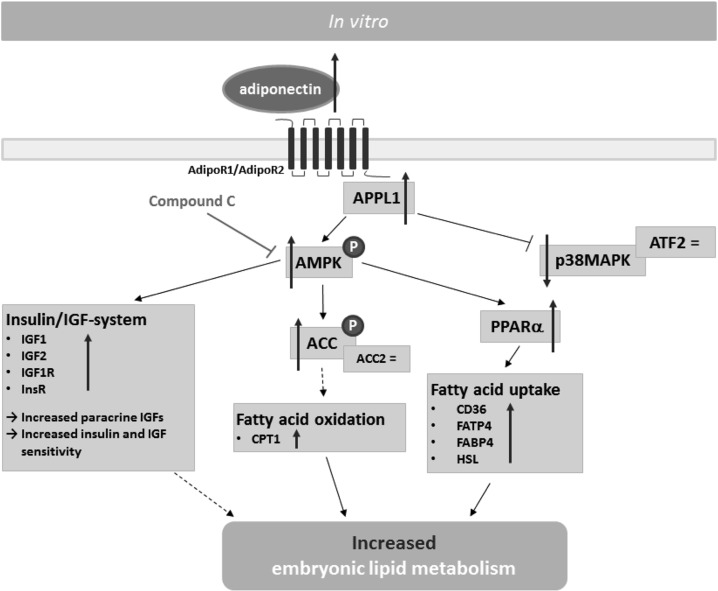
 Direct activation,
Direct activation,  Direct inhibition,
Direct inhibition,  Indirect regulation, = no change,
Indirect regulation, = no change,  increase).
increase).References
-
- Abe H, Yamashita S, Satoh T, Hoshi H. Accumulation of cytoplasmic lipid droplets in bovine embryos and cryotolerance of embryos developed in different culture systems using serum-free or serum-containing media. Mol Reprod Dev 2002;61:57–66. doi:10.1002/mrd.1131. - DOI - PubMed
-
- Aouadi M, Laurent K, Prot M, Le Marchand-Brustel Y, Binétruy B, Bost F. Inhibition of p38MAPK increases adipogenesis from embryonic to adult stages. Diabetes 2006;55:281–289. - PubMed
-
- Awazawa M, Ueki K, Inabe K, Yamauchi T, Kaneko K, Okazaki Y, Bardeesy N, Ohnishi S, Nagai R, Kadowaki T. Adiponectin suppresses hepatic SREBP1c expression in an AdipoR1/LKB1/AMPK dependent pathway. Biochem Biophys Res Commun 2009;382:51–56. doi:10.1016/j.bbrc.2009.02.131. - DOI - PubMed
-
- Bell C, Watson AJ. p38 MAPK regulates cavitation and tight junction function in the mouse blastocyst. PLOS ONE 2013;8:e59528 doi:10.1371/journal.pone.0059528. - DOI - PMC - PubMed
-
- Brookheart RT, Michel CI, Schaffer JE. As a matter of fat. Cell Metab 2009;10:9–12. doi:10.1016/j.cmet.2009.03.011. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
