

CD44high alveolar type II cells show stem cell properties during steady-state alveolar homeostasis
- PMID: 28473330
- PMCID: PMC5538873
- DOI: 10.1152/ajplung.00564.2016
CD44high alveolar type II cells show stem cell properties during steady-state alveolar homeostasis
Abstract
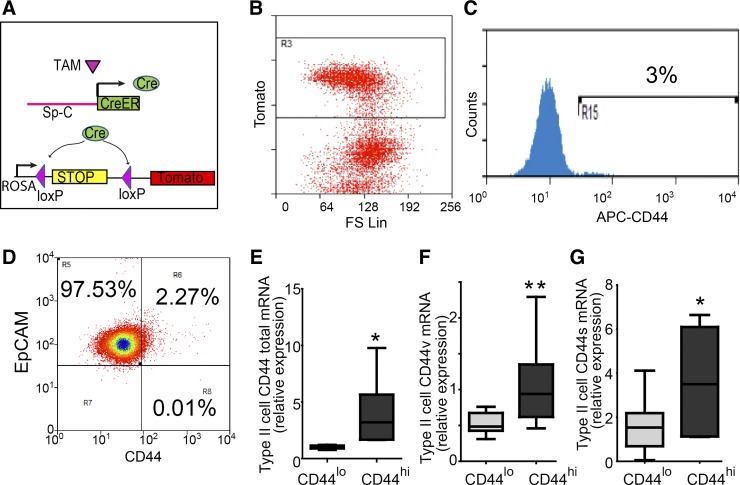
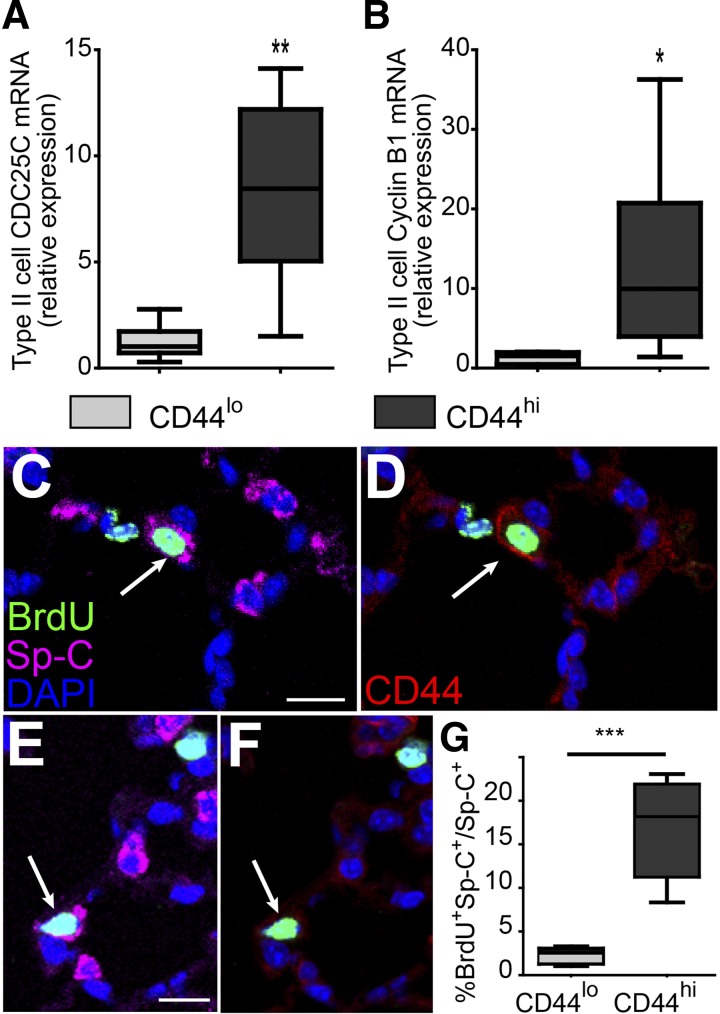
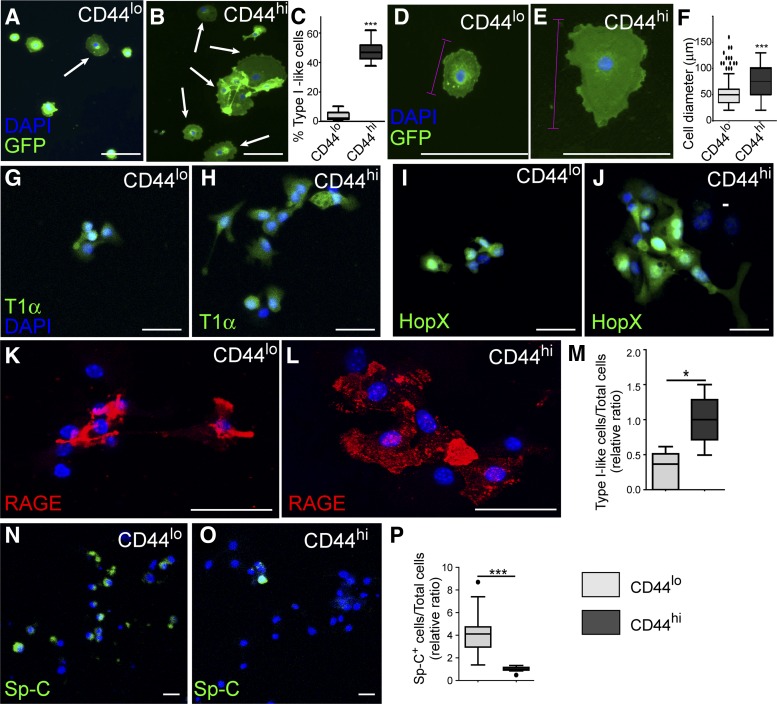
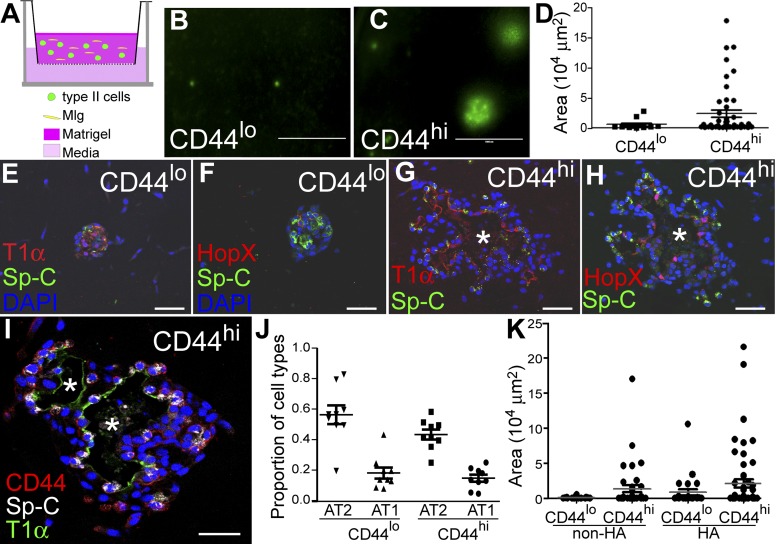
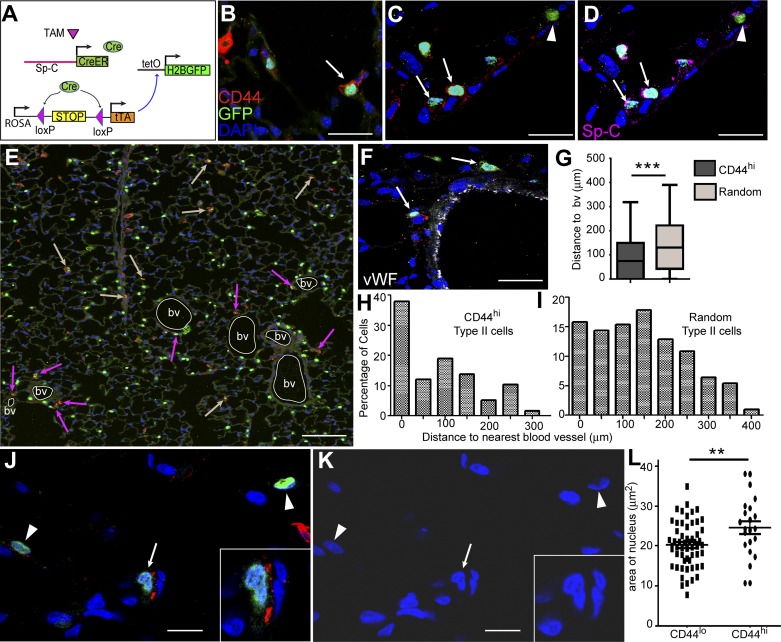
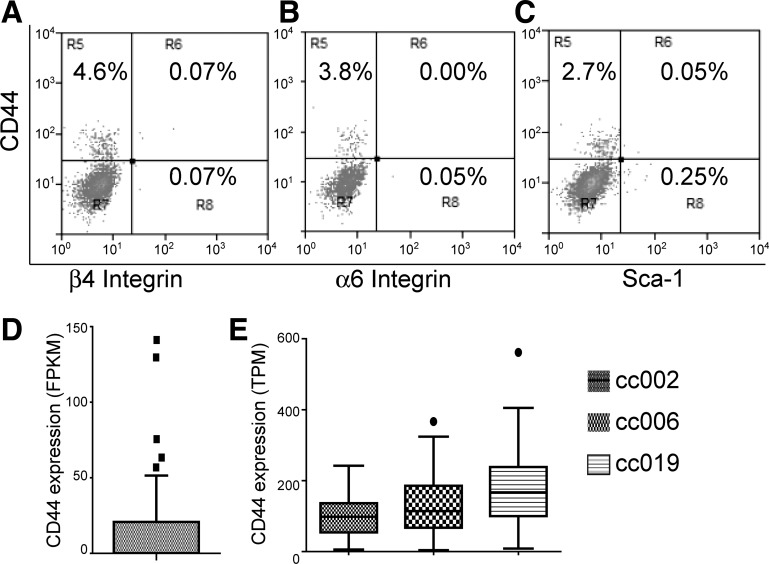
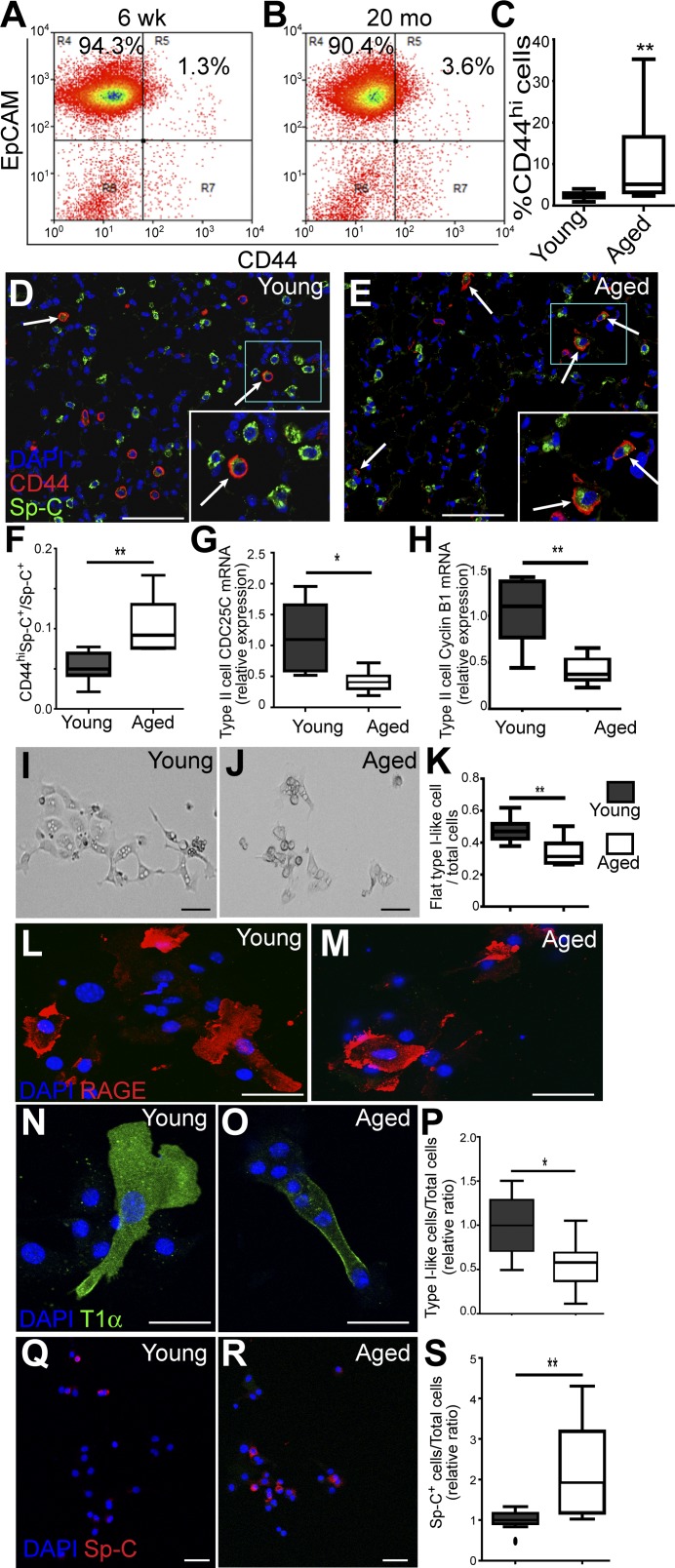
The alveolar epithelium is composed of type I cells covering most of the gas-blood exchange surface and type II cells secreting surfactant that lowers surface tension of alveoli to prevent alveolar collapse. Here, we have identified a subgroup of type II cells expressing a higher level of cell surface molecule CD44 (CD44high type II cells) that composed ~3% of total type II cells in 5-10-wk-old mice. These cells were preferentially apposed to lung capillaries. They displayed a higher proliferation rate and augmented differentiation capacity into type I cells and the ability to form alveolar organoids compared with CD44low type II cells. Moreover, in aged mice, 18-24 mo old, the percentage of CD44high type II cells among all type II cells was increased, but these cells showed decreased progenitor properties. Thus CD44high type II cells likely represent a type II cell subpopulation important for constitutive regulation of alveolar homeostasis.
Keywords: CD44; alveoli; homeostasis; lung; type II cells.
Copyright © 2017 the American Physiological Society.
Figures
References
-
- Dobbs LG. Isolation and culture of alveolar type II cells. Am J Physiol Lung Cell Mol Physiol 258: L134–L147, 1990. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous