

Aggregation of thrombin-derived C-terminal fragments as a previously undisclosed host defense mechanism
- PMID: 28473418
- PMCID: PMC5448181
- DOI: 10.1073/pnas.1619609114
Aggregation of thrombin-derived C-terminal fragments as a previously undisclosed host defense mechanism
Abstract
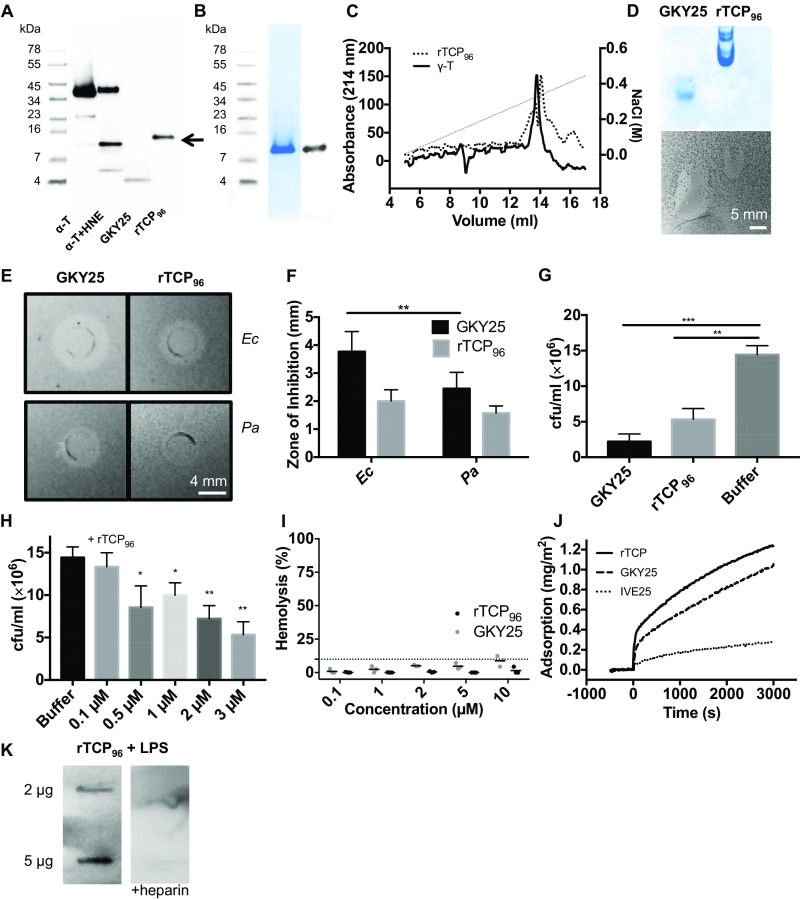
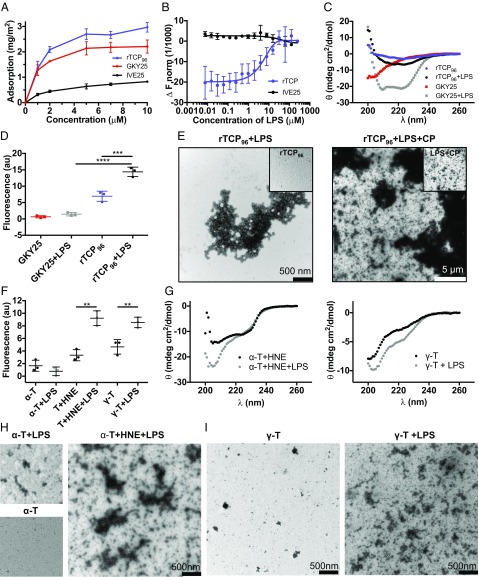
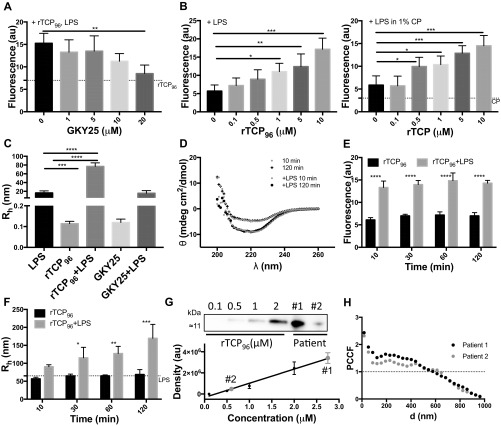
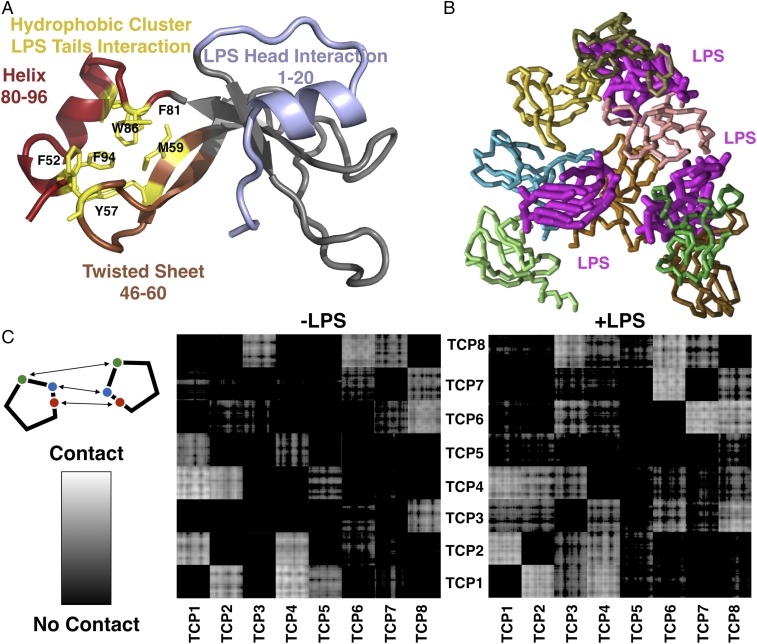
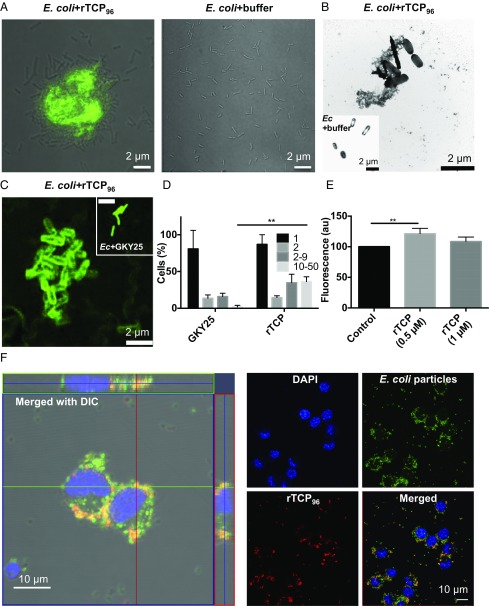
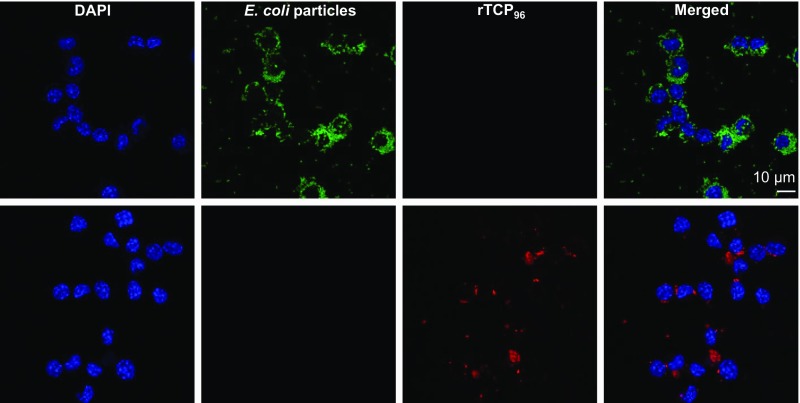
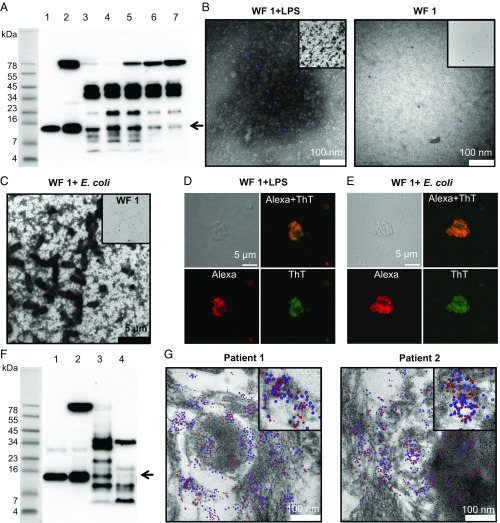
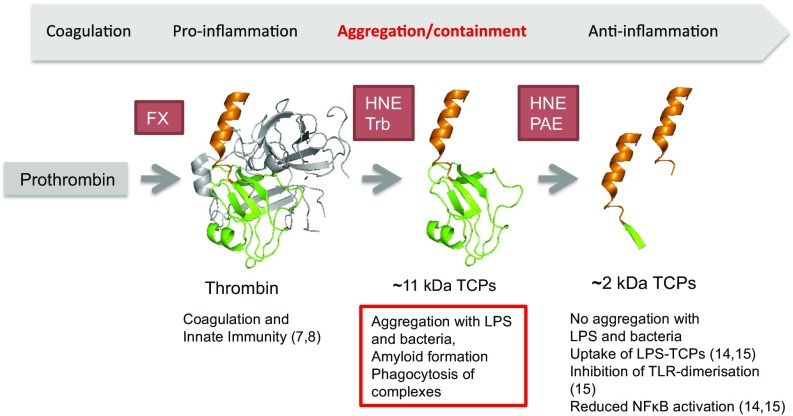
Effective control of endotoxins and bacteria is crucial for normal wound healing. During injury, the key enzyme thrombin is formed, leading to generation of fibrin. Here, we show that human neutrophil elastase cleaves thrombin, generating 11-kDa thrombin-derived C-terminal peptides (TCPs), which bind to and form amorphous amyloid-like aggregates with both bacterial lipopolysaccharide (LPS) and gram-negative bacteria. In silico molecular modeling using atomic resolution and coarse-grained simulations corroborates our experimental observations, altogether indicating increased aggregation through LPS-mediated intermolecular contacts between clusters of TCP molecules. Upon bacterial aggregation, recombinantly produced TCPs induce permeabilization of Escherichia coli and phagocytic uptake. TCPs of about 11 kDa are present in acute wound fluids as well as in fibrin sloughs from patients with infected wounds. We noted aggregation and colocalization of LPS with TCPs in such fibrin material, which indicates the presence of TCP-LPS aggregates under physiological conditions. Apart from identifying a function of proteolyzed thrombin and its fragments, our findings provide an interesting link between the coagulation system, innate immunity, LPS scavenging, and protein aggregation/amyloid formation.
Keywords: aggregation; host defense peptides; lipopolysaccharides; thrombin.
Conflict of interest statement
Conflict of interest statement: A.S. is a founder of in2cure AB, a company developing therapies based on thrombin-derived host defense peptides. The peptide GKY25 and variants are patent-protected.
Figures

Similar articles
-
Thrombin-derived C-terminal fragments aggregate and scavenge bacteria and their proinflammatory products.J Biol Chem. 2020 Mar 13;295(11):3417-3430. doi: 10.1074/jbc.RA120.012741. Epub 2020 Feb 7. J Biol Chem. 2020. PMID: 32034093 Free PMC article.
-
Structural basis for endotoxin neutralisation and anti-inflammatory activity of thrombin-derived C-terminal peptides.Nat Commun. 2018 Jul 17;9(1):2762. doi: 10.1038/s41467-018-05242-0. Nat Commun. 2018. PMID: 30018388 Free PMC article.
-
Proteolysis of human thrombin generates novel host defense peptides.PLoS Pathog. 2010 Apr 22;6(4):e1000857. doi: 10.1371/journal.ppat.1000857. PLoS Pathog. 2010. PMID: 20421939 Free PMC article.
-
Multiscale modeling of innate immune receptors: Endotoxin recognition and regulation by host defense peptides.Pharmacol Res. 2019 Sep;147:104372. doi: 10.1016/j.phrs.2019.104372. Epub 2019 Jul 24. Pharmacol Res. 2019. PMID: 31351116 Review.
-
Porphyromonas gingivalis lipopolysaccharide: an unusual pattern recognition receptor ligand for the innate host defense system.Acta Odontol Scand. 2001 Jun;59(3):131-8. doi: 10.1080/000163501750266710. Acta Odontol Scand. 2001. PMID: 11501881 Review.
Cited by
-
Cell-Free DNA Promotes Thrombin Autolysis and Generation of Thrombin-Derived C-Terminal Fragments.Front Immunol. 2021 Feb 24;12:593020. doi: 10.3389/fimmu.2021.593020. eCollection 2021. Front Immunol. 2021. PMID: 33717072 Free PMC article.
-
Antibacterial and Anti-Inflammatory Effects of Apolipoprotein E.Biomedicines. 2022 Jun 17;10(6):1430. doi: 10.3390/biomedicines10061430. Biomedicines. 2022. PMID: 35740451 Free PMC article.
-
Differential Internalization of Thrombin-Derived Host Defense Peptides into Monocytes and Macrophages.J Innate Immun. 2022;14(5):418-432. doi: 10.1159/000520831. Epub 2021 Dec 22. J Innate Immun. 2022. PMID: 34937021 Free PMC article.
-
Thrombin-Derived C-Terminal Peptide Reduces Candida-Induced Inflammation and Infection In Vitro and In Vivo.Antimicrob Agents Chemother. 2021 Oct 18;65(11):e0103221. doi: 10.1128/AAC.01032-21. Epub 2021 Aug 23. Antimicrob Agents Chemother. 2021. PMID: 34424043 Free PMC article.
-
SARS-CoV-2 spike protein aggregation is triggered by bacterial lipopolysaccharide.FEBS Lett. 2022 Oct;596(19):2566-2575. doi: 10.1002/1873-3468.14490. Epub 2022 Sep 11. FEBS Lett. 2022. PMID: 36050806 Free PMC article.
References
-
- Tennessen JA. Molecular evolution of animal antimicrobial peptides: Widespread moderate positive selection. J Evol Biol. 2005;18:1387–1394. - PubMed
-
- Zanetti M. Cathelicidins, multifunctional peptides of the innate immunity. J Leukoc Biol. 2004;75:39–48. - PubMed
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–395. - PubMed
-
- Huttner KM, Bevins CL. Antimicrobial peptides as mediators of epithelial host defense. Pediatr Res. 1999;45:785–794. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
