

Modeling Renal Cell Carcinoma in Mice: Bap1 and Pbrm1 Inactivation Drive Tumor Grade
- PMID: 28473526
- PMCID: PMC5540776
- DOI: 10.1158/2159-8290.CD-17-0292
Modeling Renal Cell Carcinoma in Mice: Bap1 and Pbrm1 Inactivation Drive Tumor Grade
Abstract
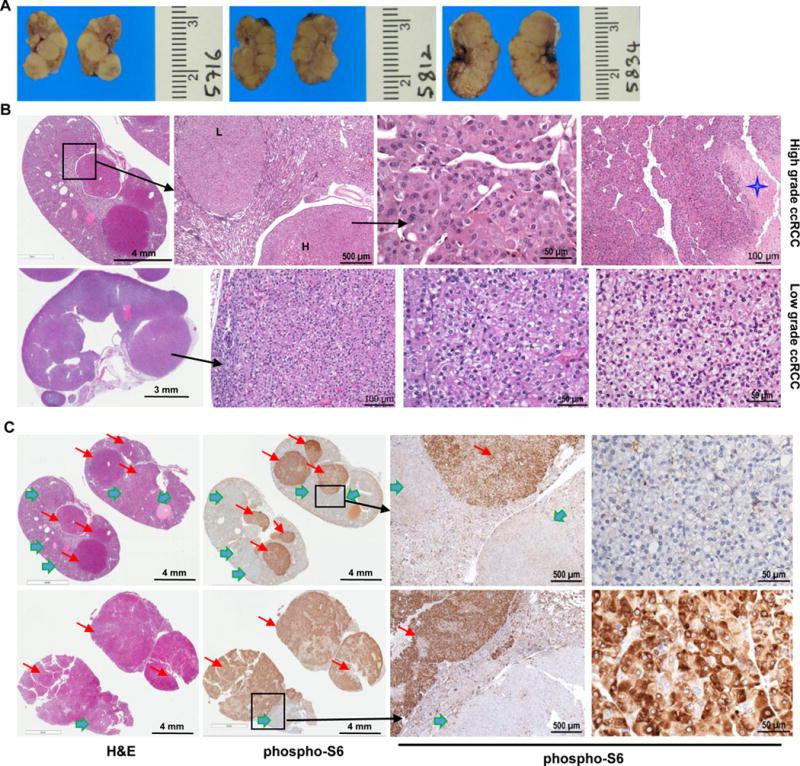
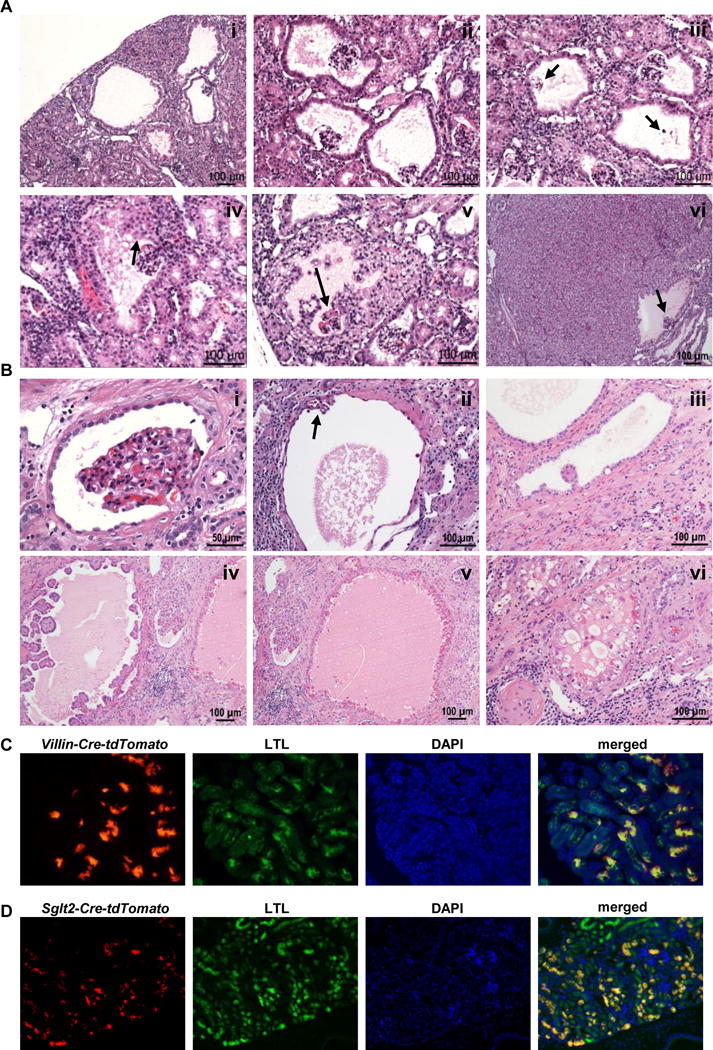
Clear cell renal cell carcinoma (ccRCC) is characterized by BAP1 and PBRM1 mutations, which are associated with tumors of different grade and prognosis. However, whether BAP1 and PBRM1 loss causes ccRCC and determines tumor grade is unclear. We conditionally targeted Bap1 and Pbrm1 (with Vhl) in the mouse using several Cre drivers. Sglt2 and Villin proximal convoluted tubule drivers failed to cause tumorigenesis, challenging the conventional notion of ccRCC origins. In contrast, targeting with PAX8, a transcription factor frequently overexpressed in ccRCC, led to ccRCC of different grades. Bap1-deficient tumors were of high grade and showed greater mTORC1 activation than Pbrm1-deficient tumors, which exhibited longer latency. Disrupting one allele of the mTORC1 negative regulator, Tsc1, in Pbrm1-deficient kidneys triggered higher grade ccRCC. This study establishes Bap1 and Pbrm1 as lineage-specific drivers of ccRCC and histologic grade, implicates mTORC1 as a tumor grade rheostat, and suggests that ccRCCs arise from Bowman capsule cells.Significance: Determinants of tumor grade and aggressiveness across cancer types are poorly understood. Using ccRCC as a model, we show that Bap1 and Pbrm1 loss drives tumor grade. Furthermore, we show that the conversion from low grade to high grade can be promoted by activation of mTORC1. Cancer Discov; 7(8); 900-17. ©2017 AACR.See related commentary by Leung and Kim, p. 802This article is highlighted in the In This Issue feature, p. 783.
©2017 American Association for Cancer Research.
Conflict of interest statement
Figures
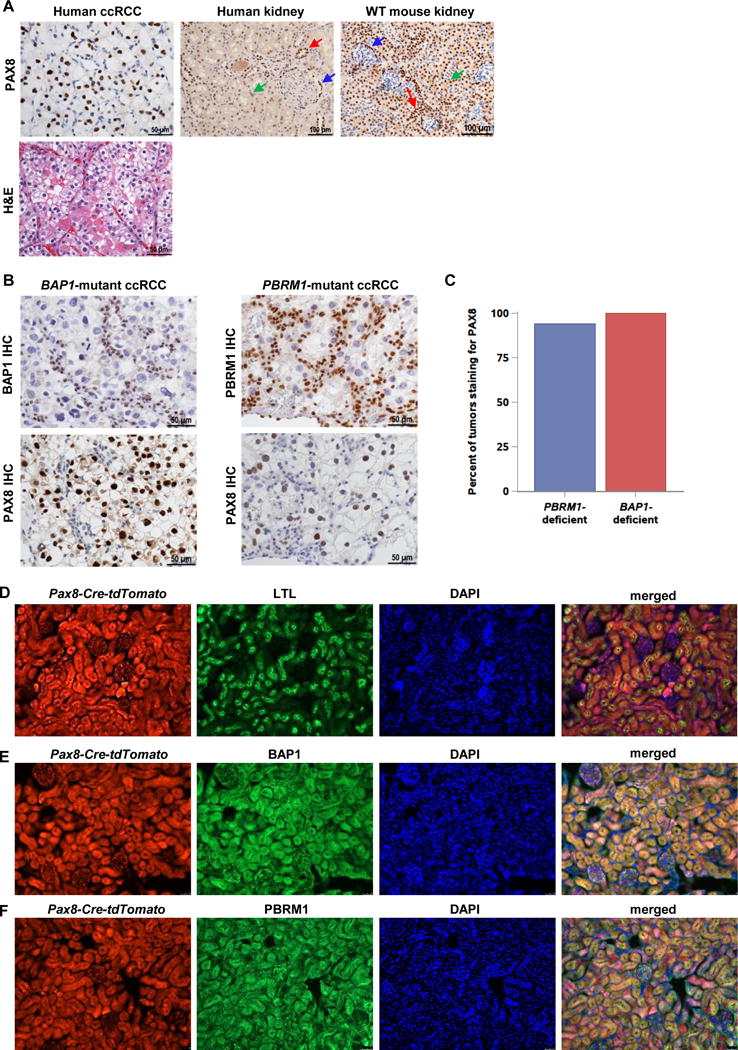
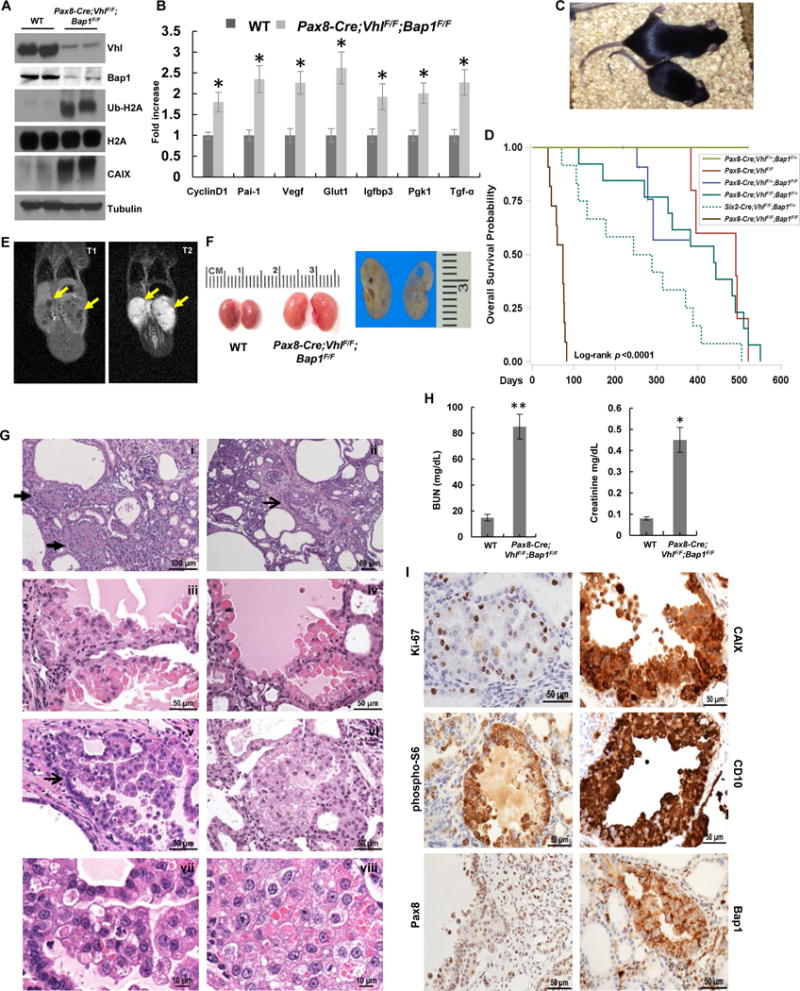
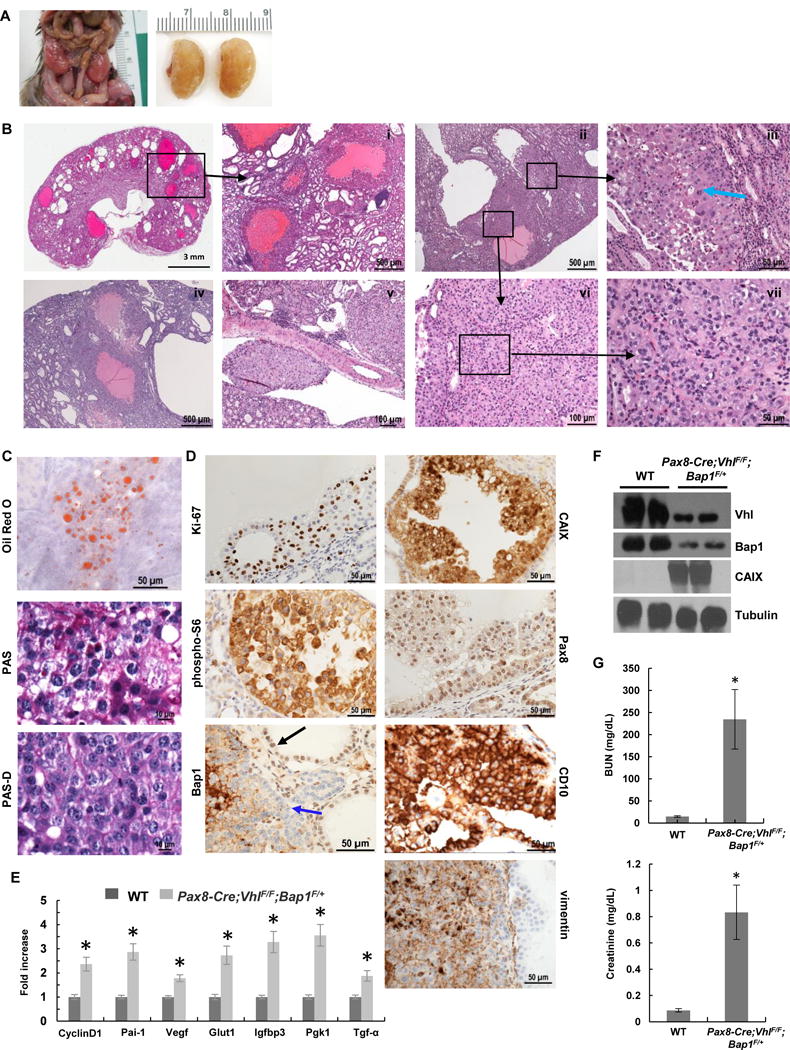
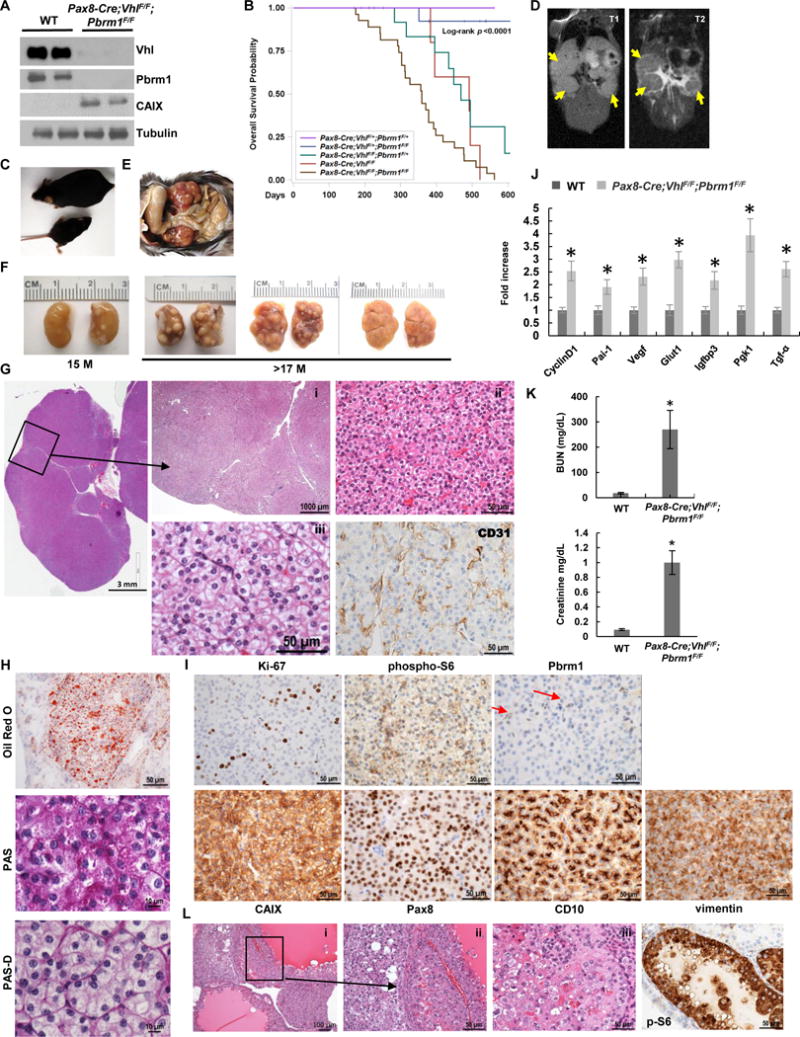
Comment in
-
Bap1 and Pbrm1: Determinants of Tumor Grade and mTOR Activation in VHL-Deficient Mouse Models of Renal Cell Carcinoma.Cancer Discov. 2017 Aug;7(8):802-804. doi: 10.1158/2159-8290.CD-17-0610. Cancer Discov. 2017. PMID: 28765116 Free PMC article.
References
-
- Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA: a cancer journal for clinicians. 2013;63(1):11–30. - PubMed
-
- Baldewijns MM, van Vlodrop IJ, Schouten LJ, Soetekouw PM, de Bruine AP, van Engeland M. Genetics and epigenetics of renal cell cancer. Biochimica et Biophysica Acta. 2008;1785(2):133–55. - PubMed
-
- Gnarra JR, Tory K, Weng Y, Schmidt L, Wei MH, Li H, Latif F, Liu S, Chen F, Duh FM, et al. Mutations of the VHL tumour suppressor gene in renal carcinoma. Nature Genetics. 1994;7(1):85–90. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
