

Branch-specific plasticity of a bifunctional dopamine circuit encodes protein hunger
- PMID: 28473588
- PMCID: PMC5513152
- DOI: 10.1126/science.aal3245
Branch-specific plasticity of a bifunctional dopamine circuit encodes protein hunger
Abstract
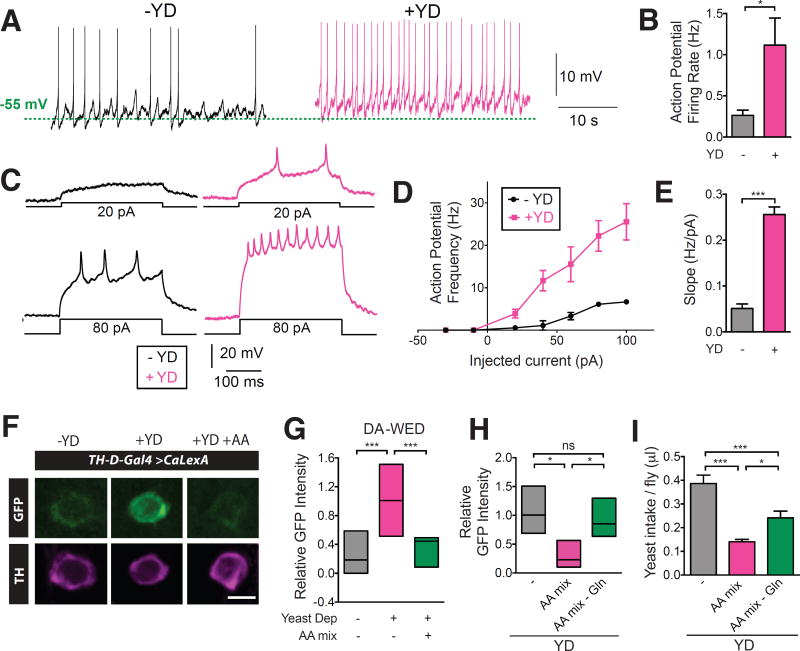
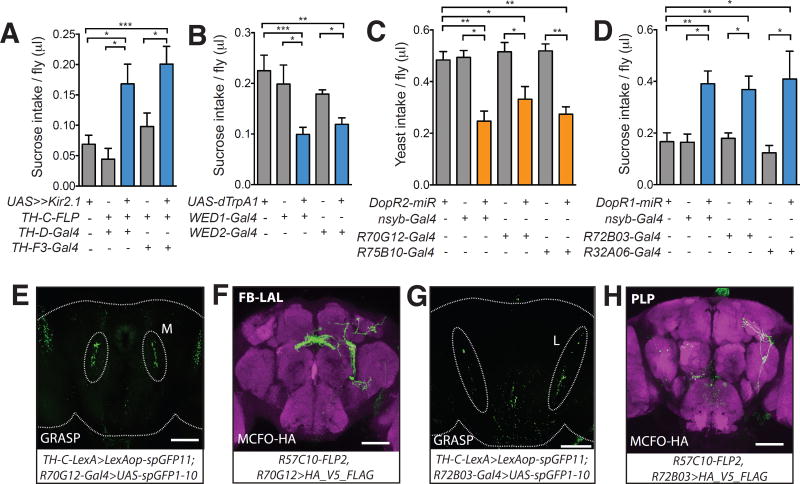
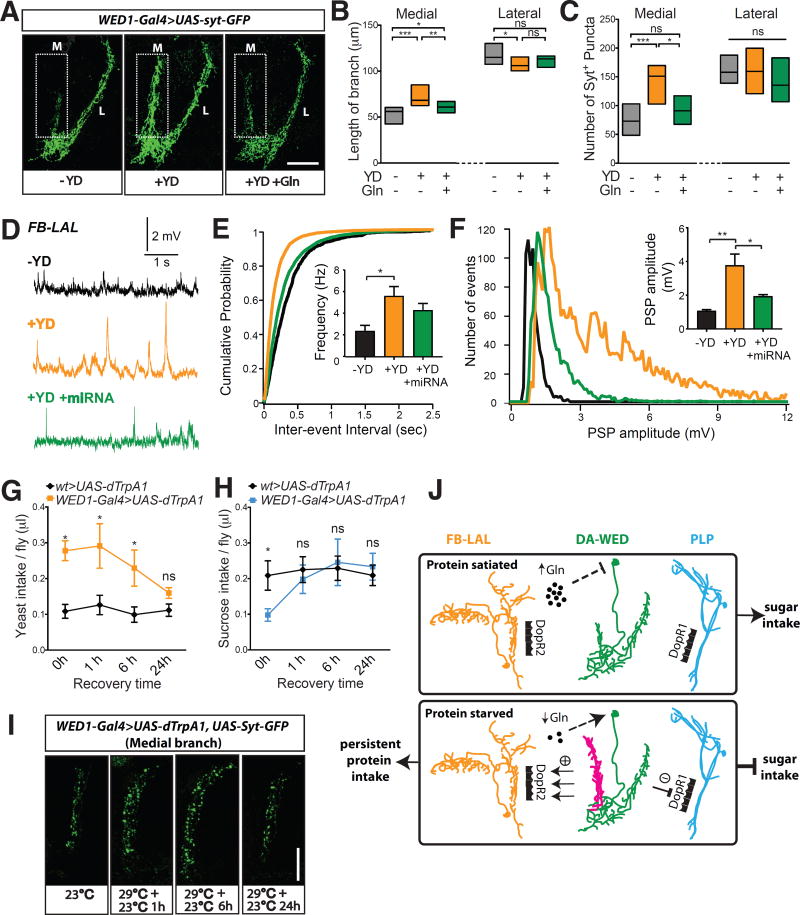
Free-living animals must not only regulate the amount of food they consume but also choose which types of food to ingest. The shifting of food preference driven by nutrient-specific hunger can be essential for survival, yet little is known about the underlying mechanisms. We identified a dopamine circuit that encodes protein-specific hunger in Drosophila The activity of these neurons increased after substantial protein deprivation. Activation of this circuit simultaneously promoted protein intake and restricted sugar consumption, via signaling to distinct downstream neurons. Protein starvation triggered branch-specific plastic changes in these dopaminergic neurons, thus enabling sustained protein consumption. These studies reveal a crucial circuit mechanism by which animals adjust their dietary strategy to maintain protein homeostasis.
Copyright © 2017, American Association for the Advancement of Science.
Figures

References
-
- Richter CP. Increased salt appetite in adrenalectomized rats. American Journal of Physiology. 1936;115:155–161.
-
- Richter CP. Total self regulatory functions of animals and human beings. (Harvey Lecture Series, 1942–1943) 38
-
- Simpson SJ, Raubenbeimer D. The Nature of Nutrition. Princeton University Press; 2012.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
