

FnrL and Three Dnr Regulators Are Used for the Metabolic Adaptation to Low Oxygen Tension in Dinoroseobacter shibae
- PMID: 28473807
- PMCID: PMC5398030
- DOI: 10.3389/fmicb.2017.00642
FnrL and Three Dnr Regulators Are Used for the Metabolic Adaptation to Low Oxygen Tension in Dinoroseobacter shibae
Abstract
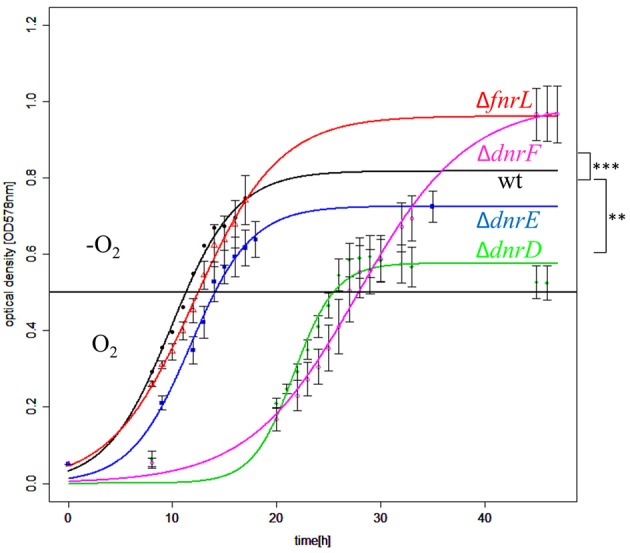
The heterotrophic marine bacterium Dinoroseobacter shibae utilizes aerobic respiration and anaerobic denitrification supplemented with aerobic anoxygenic photosynthesis for energy generation. The aerobic to anaerobic transition is controlled by four Fnr/Crp family regulators in a unique cascade-type regulatory network. FnrL is utilizing an oxygen-sensitive Fe-S cluster for oxygen sensing. Active FnrL is inducing most operons encoding the denitrification machinery and the corresponding heme biosynthesis. Activation of gene expression of the high oxygen affinity cbb3-type and repression of the low affinity aa3-type cytochrome c oxidase is mediated by FnrL. Five regulator genes including dnrE and dnrF are directly controlled by FnrL. Multiple genes of the universal stress protein (USP) and cold shock response are further FnrL targets. DnrD, most likely sensing NO via a heme cofactor, co-induces genes of denitrification, heme biosynthesis, and the regulator genes dnrE and dnrF. DnrE is controlling genes for a putative Na+/H+ antiporter, indicating a potential role of a Na+ gradient under anaerobic conditions. The formation of the electron donating primary dehydrogenases is coordinated by FnrL and DnrE. Many plasmid encoded genes were DnrE regulated. DnrF is controlling directly two regulator genes including the Fe-S cluster biosynthesis regulator iscR, genes of the electron transport chain and the glutathione metabolism. The genes for nitrate reductase and CO dehydrogenase are repressed by DnrD and DnrF. Both regulators in concert with FnrL are inducing the photosynthesis genes. One of the major denitrification operon control regions, the intergenic region between nirS and nosR2, contains one Fnr/Dnr binding site. Using regulator gene mutant strains, lacZ-reporter gene fusions in combination with promoter mutagenesis, the function of the single Fnr/Dnr binding site for FnrL-, DnrD-, and partly DnrF-dependent nirS and nosR2 transcriptional activation was shown. Overall, the unique regulatory network of the marine bacterium D. shibae for the transition from aerobic to anaerobic growth composed of four Crp/Fnr family regulators was elucidated.
Keywords: Crp/Fnr regulator; Dinoroseobacter shibae; Dnr; FnrL; anaerobic energy metabolism; denitrification; oxygen-dependent gene expression; regulation.
Figures









References
-
- Becher A., Schweizer H. P. (2000). Integration-proficient Pseudomonas aeruginosa vectors for isolation of single-copy chromosomal lacZ and lux gene fusions. Biotechniques 29, 948–952. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous