

Stemness in Cancer: Stem Cells, Cancer Stem Cells, and Their Microenvironment
- PMID: 28473858
- PMCID: PMC5394399
- DOI: 10.1155/2017/5619472
Stemness in Cancer: Stem Cells, Cancer Stem Cells, and Their Microenvironment
Abstract
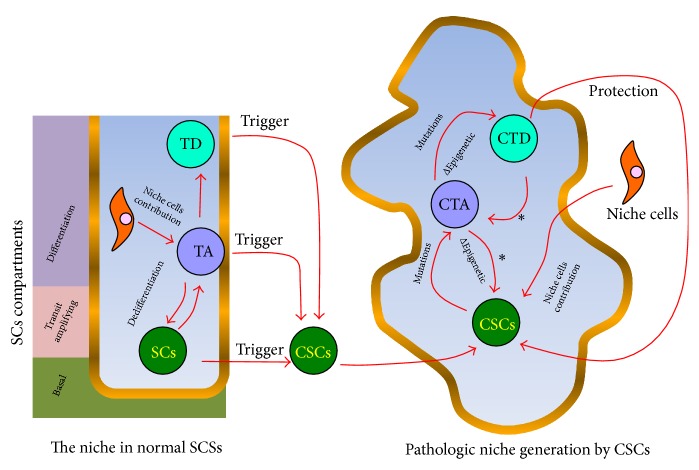
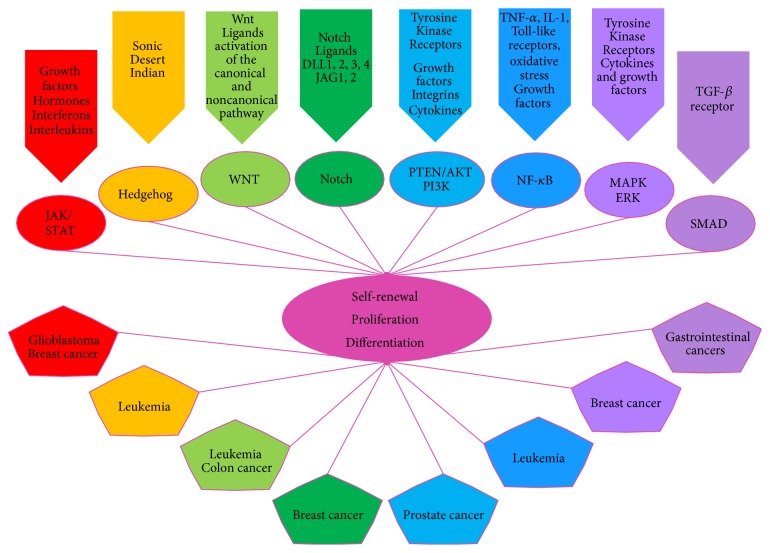
Stemness combines the ability of a cell to perpetuate its lineage, to give rise to differentiated cells, and to interact with its environment to maintain a balance between quiescence, proliferation, and regeneration. While adult Stem Cells display these properties when participating in tissue homeostasis, Cancer Stem Cells (CSCs) behave as their malignant equivalents. CSCs display stemness in various circumstances, including the sustaining of cancer progression, and the interaction with their environment in search for key survival factors. As a result, CSCs can recurrently persist after therapy. In order to understand how the concept of stemness applies to cancer, this review will explore properties shared between normal and malignant Stem Cells. First, we provide an overview of properties of normal adult Stem Cells. We thereafter elaborate on how these features operate in CSCs. We then review the organization of microenvironment components, which enables CSCs hosting. We subsequently discuss Mesenchymal Stem/Stromal Cells (MSCs), which, although their stemness properties are limited, represent essential components of the Stem Cell niche and tumor microenvironment. We next provide insights of the therapeutic strategies targeting Stem Cell properties in tumors and the use of state-of-the-art techniques in future research. Increasing our knowledge of the CSCs microenvironment is key to identifying new therapeutic solutions.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
