

Complete Freund's adjuvant induces experimental autoimmune myocarditis by enhancing IL-6 production during initiation of the immune response
- PMID: 28474508
- PMCID: PMC5418134
- DOI: 10.1002/iid3.155
Complete Freund's adjuvant induces experimental autoimmune myocarditis by enhancing IL-6 production during initiation of the immune response
Abstract
Introduction: Complete Freund's Adjuvant (CFA) emulsified with an antigen is a widely used method to induce autoimmune disease in animal models, yet the contribution of CFA to the immune response is not well understood. We compared the effectiveness of CFA with Incomplete Freund's Adjuvant (IFA) or TiterMax Gold Adjuvant (TMax) in experimental autoimmune myocarditis (EAM) in male mice.
Methods: EAM was induced in A/J, BALB/c, and IL6KO BALB/c male mice by injection of the myocarditogenic peptide in CFA, IFA, or TMax on days 0 and 7. EAM severity was analyzed by histology on day 21. In addition, specific flow cytometry outcomes were evaluated on day 21.
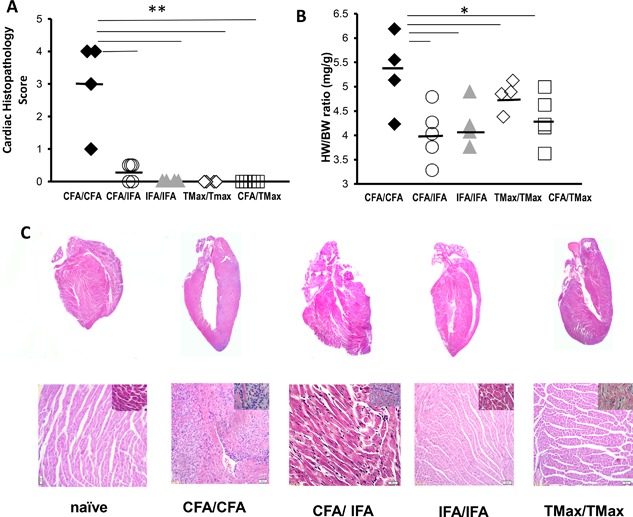
Results: Only mice immunized with CFA and myocarditogenic peptide on both days 0 and 7 developed substantial myocarditis as measured by histology. We observed a significantly increased level of IL6 in the spleen 3 days after CFA immunization. In the spleen and heart on day 21, there was an expansion of myeloid cells in CFA-immunized mice, as compared to IFA or TMax-immunized animals. Recombinant IL-6 at the time of IFA immunization partially restored susceptibility of the mice to EAM. We also treated EAM-resistant IL-6 knockout mice with recombinant IL-6 around the time of the first immunization, on days -1 to 2, completely restoring disease susceptibility, showing that the requirement for IL-6 coincides with primary immunization. Examining APC populations in the lymph node draining the immunization site evidenced the contribution of IL-6 to the CFA-dependence of EAM was through controlling local dendritic cell (DC) trafficking.
Conclusions: CFA used with myocarditogenic peptide twice is required to induce EAM in both A/J and Balb/c mice. Although IFA and TiterMax induce antibody responses, only CFA preferentially induced autoantigen-specific responses. CFA expands monocytes in the heart and in the spleen. IL-6 signaling is required during short window around primary immunization to induce EAM. In addition, IL-6 deficient mice resistance to EAM could be reversed by injecting IL-6 around first immunization. IL-6 expands dendritic cell and monocytic populations and ultimately leads to a robust T-cell driven immune response in CFA immunized mice.
Keywords: Autoimmunity; adjuvant; myocarditis.
© 2017 The Authors. Immunity, Inflammation and Disease Published by John Wiley & Sons Ltd.
Figures






Similar articles
-
Prominin-1+/CD133+ bone marrow-derived heart-resident cells suppress experimental autoimmune myocarditis.Cardiovasc Res. 2008 Nov 1;80(2):236-45. doi: 10.1093/cvr/cvn190. Epub 2008 Jul 11. Cardiovasc Res. 2008. PMID: 18621802
-
An alternative technique for the induction of autoimmune valvulitis in a rat model of rheumatic heart disease.J Immunol Methods. 2010 Apr 15;355(1-2):80-5. doi: 10.1016/j.jim.2010.02.013. Epub 2010 Mar 3. J Immunol Methods. 2010. PMID: 20206182
-
GM-CSF promotes inflammatory dendritic cell formation but does not contribute to disease progression in experimental autoimmune myocarditis.Biochim Biophys Acta. 2013 Apr;1833(4):934-44. doi: 10.1016/j.bbamcr.2012.10.008. Epub 2012 Oct 24. Biochim Biophys Acta. 2013. PMID: 23103516
-
Myocarditogenic epitopes and autoimmune myocarditis.Intern Med. 2003 Jan;42(1):3-6. doi: 10.2169/internalmedicine.42.3. Intern Med. 2003. PMID: 12583610 Review.
-
Complete Freund's adjuvant as a confounding factor in multiple sclerosis research.Front Immunol. 2024 Feb 15;15:1353865. doi: 10.3389/fimmu.2024.1353865. eCollection 2024. Front Immunol. 2024. PMID: 38426111 Free PMC article. Review.
Cited by
-
The Outer Membrane Proteins and Their Synergy Triggered the Protective Effects against Pathogenic Escherichia coli.Microorganisms. 2022 May 8;10(5):982. doi: 10.3390/microorganisms10050982. Microorganisms. 2022. PMID: 35630426 Free PMC article.
-
A comprehensive review on anti-inflammatory plants: a mechanistic insight through preclinical and clinical studies.Inflammopharmacology. 2025 May;33(5):2447-2476. doi: 10.1007/s10787-025-01764-4. Epub 2025 Jun 4. Inflammopharmacology. 2025. PMID: 40465037 Review.
-
A juvenile mouse model of anti-N-methyl-D-aspartate receptor encephalitis by active immunization.Front Mol Neurosci. 2023 Sep 18;16:1211119. doi: 10.3389/fnmol.2023.1211119. eCollection 2023. Front Mol Neurosci. 2023. PMID: 37790883 Free PMC article.
-
Metabolic Reprogramming in Response to Freund's Adjuvants: Insights from Serum Metabolomics.Microorganisms. 2025 Feb 22;13(3):492. doi: 10.3390/microorganisms13030492. Microorganisms. 2025. PMID: 40142385 Free PMC article.
-
Transcriptomic Analysis of Inflammatory Cardiomyopathy Identifies Molecular Signatures of Disease and Informs in silico Prediction of a Network-Based Rationale for Therapy.Front Immunol. 2021 Mar 5;12:640837. doi: 10.3389/fimmu.2021.640837. eCollection 2021. Front Immunol. 2021. PMID: 33746983 Free PMC article.
References
-
- Petrovsky, N. , and Aguilar J. C.. 2004. Vaccine adjuvants: current state and future trends. Immunol. Cell Biol. 82:488–496. - PubMed
-
- Singh, M. , and O'Hagan D. T.. 2002. Recent advances in vaccine adjuvants. Pharm. Res. 19:715–728. - PubMed
-
- Chang, S. P. , Nikaido C. M., Hashimoto A. C., Hashiro C. Q., Yokota B. T., and Hui G. S.. 1994. Regulation of antibody specificity to Plasmodium falciparum merozoite surface protein‐1 by adjuvant and MHC haplotype. J. Immunol. 152:3483–3490. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
