

The molecular mechanism of the type IVa pilus motors
- PMID: 28474682
- PMCID: PMC5424180
- DOI: 10.1038/ncomms15091
The molecular mechanism of the type IVa pilus motors
Abstract
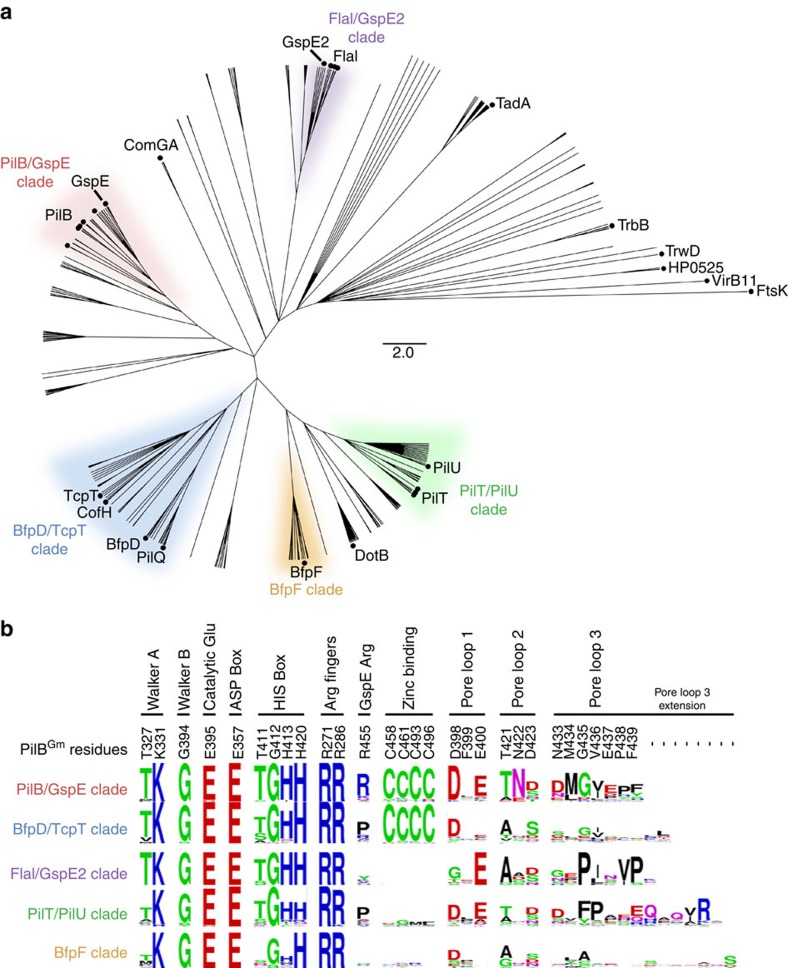
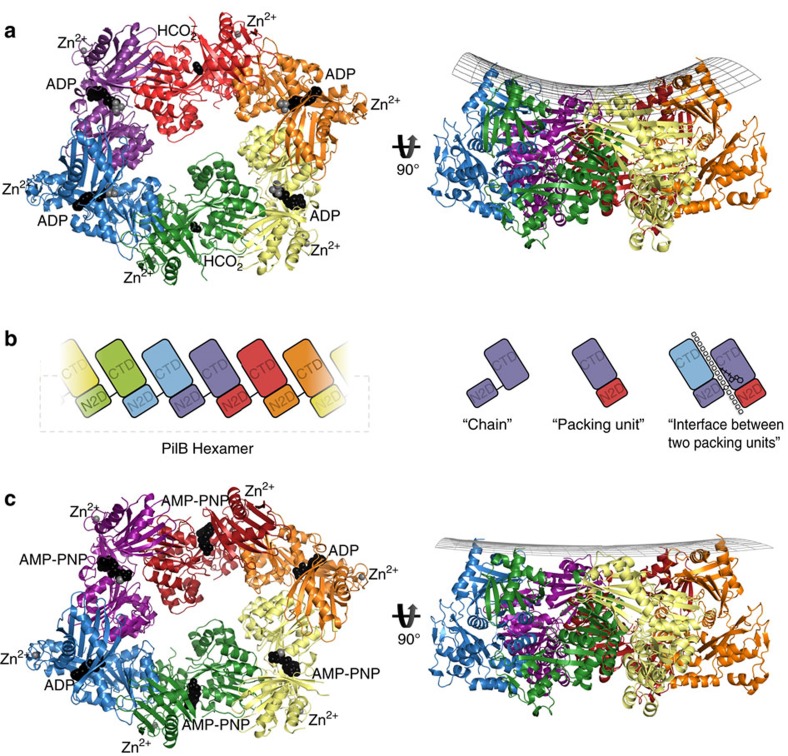
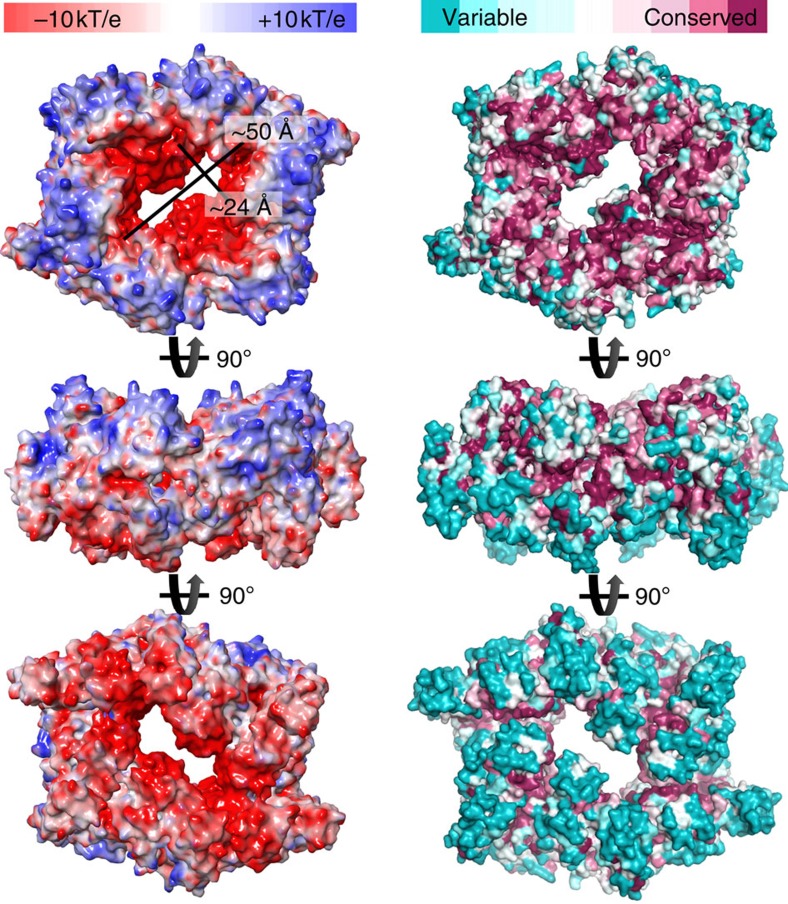
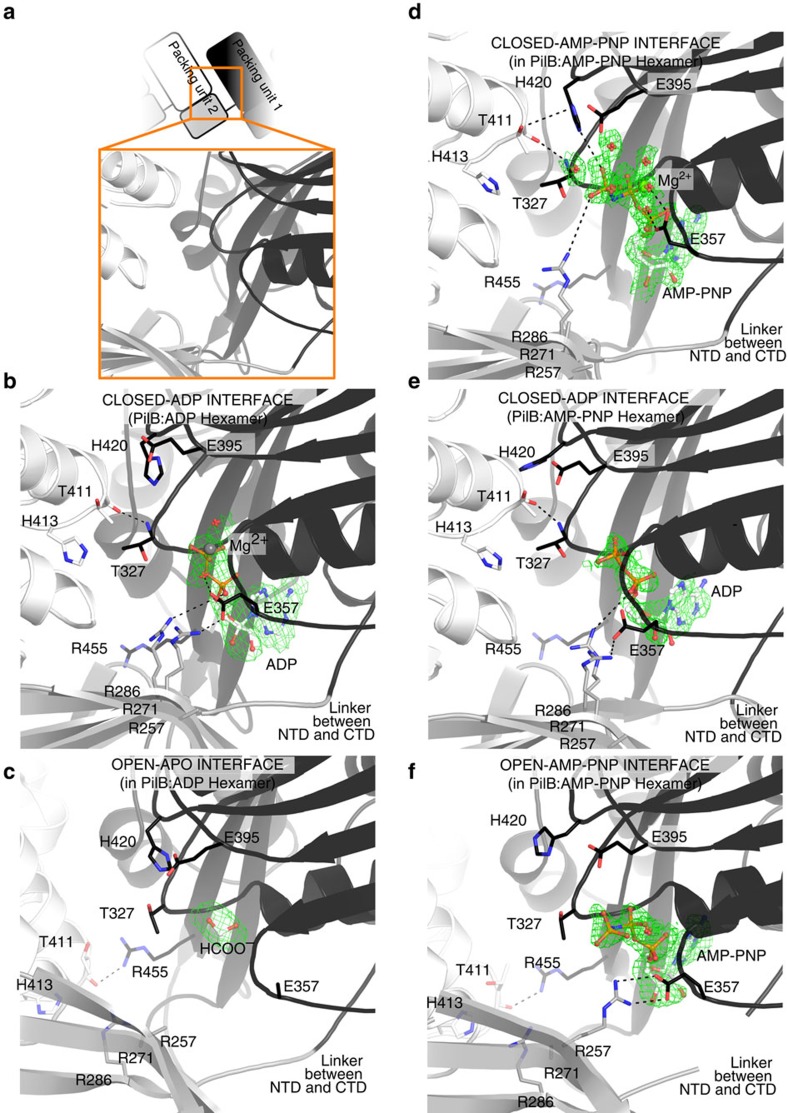
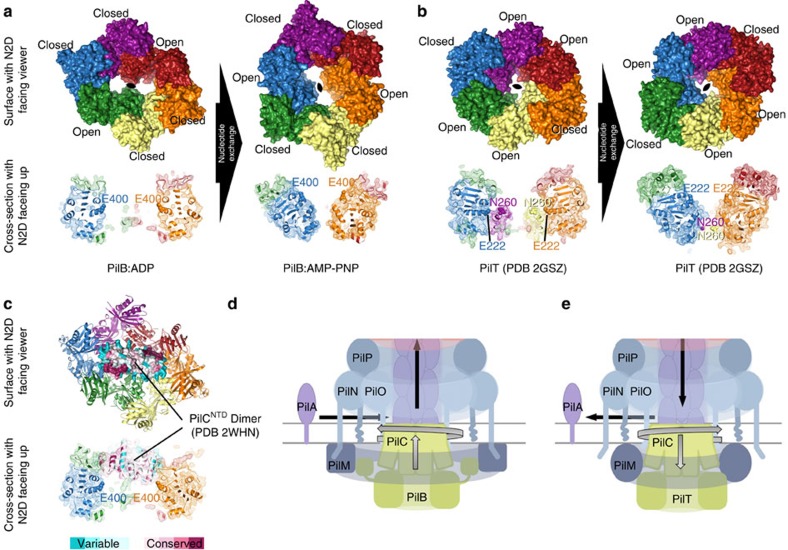
Type IVa pili are protein filaments essential for virulence in many bacterial pathogens; they extend and retract from the surface of bacterial cells to pull the bacteria forward. The motor ATPase PilB powers pilus assembly. Here we report the structures of the core ATPase domains of Geobacter metallireducens PilB bound to ADP and the non-hydrolysable ATP analogue, AMP-PNP, at 3.4 and 2.3 Å resolution, respectively. These structures reveal important differences in nucleotide binding between chains. Analysis of these differences reveals the sequential turnover of nucleotide, and the corresponding domain movements. Our data suggest a clockwise rotation of the central sub-pores of PilB, which through interactions with PilC, would support the assembly of a right-handed helical pilus. Our analysis also suggests a counterclockwise rotation of the C2 symmetric PilT that would enable right-handed pilus disassembly. The proposed model provides insight into how this family of ATPases can power pilus extension and retraction.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Structural insights into the mechanism of Type IVa pilus extension and retraction ATPase motors.FEBS J. 2018 Sep;285(18):3402-3421. doi: 10.1111/febs.14619. Epub 2018 Aug 18. FEBS J. 2018. PMID: 30066435
-
Retraction ATPase Motors from Three Orthologous Type IVa Pilus Systems Support Promiscuous Retraction of the Vibrio cholerae Competence Pilus.J Bacteriol. 2022 Jun 21;204(6):e0012622. doi: 10.1128/jb.00126-22. Epub 2022 May 4. J Bacteriol. 2022. PMID: 35506694 Free PMC article.
-
The Type IV Pilus Assembly ATPase PilB of Myxococcus xanthus Interacts with the Inner Membrane Platform Protein PilC and the Nucleotide-binding Protein PilM.J Biol Chem. 2016 Mar 25;291(13):6946-57. doi: 10.1074/jbc.M115.701284. Epub 2016 Feb 5. J Biol Chem. 2016. PMID: 26851283 Free PMC article.
-
Building permits-control of type IV pilus assembly by PilB and its cofactors.J Bacteriol. 2024 Dec 19;206(12):e0035924. doi: 10.1128/jb.00359-24. Epub 2024 Nov 7. J Bacteriol. 2024. PMID: 39508682 Free PMC article. Review.
-
Pulling together with type IV pili.J Mol Microbiol Biotechnol. 2004;7(1-2):52-62. doi: 10.1159/000077869. J Mol Microbiol Biotechnol. 2004. PMID: 15170403 Review.
Cited by
-
Motile ghosts of the halophilic archaeon, Haloferax volcanii.Proc Natl Acad Sci U S A. 2020 Oct 27;117(43):26766-26772. doi: 10.1073/pnas.2009814117. Epub 2020 Oct 13. Proc Natl Acad Sci U S A. 2020. PMID: 33051299 Free PMC article.
-
More than a feeling: microscopy approaches to understanding surface-sensing mechanisms.J Bacteriol. 2020 Oct 19;203(6):e00492-20. doi: 10.1128/JB.00492-20. Online ahead of print. J Bacteriol. 2020. PMID: 33077631 Free PMC article. Review.
-
PilT and PilU are homohexameric ATPases that coordinate to retract type IVa pili.PLoS Genet. 2019 Oct 18;15(10):e1008448. doi: 10.1371/journal.pgen.1008448. eCollection 2019 Oct. PLoS Genet. 2019. PMID: 31626631 Free PMC article.
-
Decoupling Filamentous Phage Uptake and Energy of the TolQRA Motor in Escherichia coli.J Bacteriol. 2020 Jan 2;202(2):e00428-19. doi: 10.1128/JB.00428-19. Print 2020 Jan 2. J Bacteriol. 2020. PMID: 31636109 Free PMC article.
-
Mechanism of assembly of type 4 filaments: everything you always wanted to know (but were afraid to ask).Microbiology (Reading). 2023 Mar;169(3):001311. doi: 10.1099/mic.0.001311. Microbiology (Reading). 2023. PMID: 36947586 Free PMC article. Review.
References
-
- Burrows L. L. Pseudomonas aeruginosa twitching motility: type IV pili in action. Annu. Rev. Microbiol. 66, 493–520 (2012). - PubMed
-
- Peabody C. R. et al.. Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. Microbiology 149, 3051–3072 (2003). - PubMed
-
- Mattick J. S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 56, 289–314 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
