

Recurrent Potent Human Neutralizing Antibodies to Zika Virus in Brazil and Mexico
- PMID: 28475892
- PMCID: PMC5492969
- DOI: 10.1016/j.cell.2017.04.024
Recurrent Potent Human Neutralizing Antibodies to Zika Virus in Brazil and Mexico
Abstract
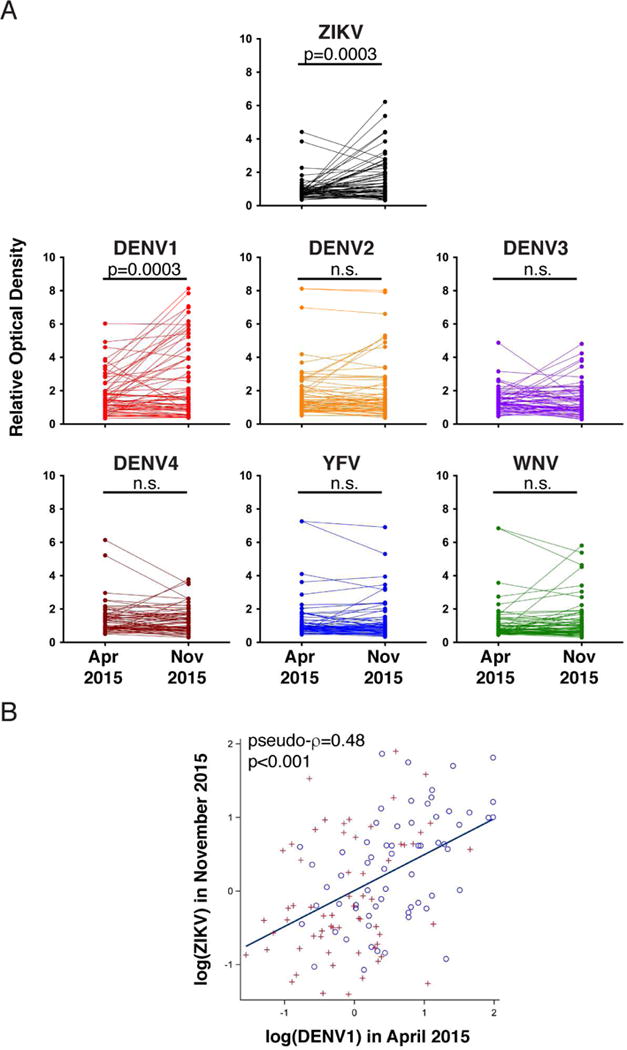
Antibodies to Zika virus (ZIKV) can be protective. To examine the antibody response in individuals who develop high titers of anti-ZIKV antibodies, we screened cohorts in Brazil and Mexico for ZIKV envelope domain III (ZEDIII) binding and neutralization. We find that serologic reactivity to dengue 1 virus (DENV1) EDIII before ZIKV exposure is associated with increased ZIKV neutralizing titers after exposure. Antibody cloning shows that donors with high ZIKV neutralizing antibody titers have expanded clones of memory B cells that express the same immunoglobulin VH3-23/VK1-5 genes. These recurring antibodies cross-react with DENV1, but not other flaviviruses, neutralize both DENV1 and ZIKV, and protect mice against ZIKV challenge. Structural analyses reveal the mechanism of recognition of the ZEDIII lateral ridge by VH3-23/VK1-5 antibodies. Serologic testing shows that antibodies to this region correlate with serum neutralizing activity to ZIKV. Thus, high neutralizing responses to ZIKV are associated with pre-existing reactivity to DENV1 in humans.
Keywords: Zika virus; antibodies; dengue virus; flavivirus; structure; vaccine.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures






Similar articles
-
Longitudinal Analysis of Antibody Cross-neutralization Following Zika Virus and Dengue Virus Infection in Asia and the Americas.J Infect Dis. 2018 Jul 13;218(4):536-545. doi: 10.1093/infdis/jiy164. J Infect Dis. 2018. PMID: 29618091 Free PMC article.
-
Immunization of Zika virus envelope protein domain III induces specific and neutralizing immune responses against Zika virus.Vaccine. 2017 Jul 24;35(33):4287-4294. doi: 10.1016/j.vaccine.2017.04.052. Epub 2017 Jun 29. Vaccine. 2017. PMID: 28669618 Free PMC article.
-
Structural basis for Zika envelope domain III recognition by a germline version of a recurrent neutralizing antibody.Proc Natl Acad Sci U S A. 2020 May 5;117(18):9865-9875. doi: 10.1073/pnas.1919269117. Epub 2020 Apr 22. Proc Natl Acad Sci U S A. 2020. PMID: 32321830 Free PMC article.
-
Recent advances in understanding the adaptive immune response to Zika virus and the effect of previous flavivirus exposure.Virus Res. 2018 Aug 2;254:27-33. doi: 10.1016/j.virusres.2017.06.019. Epub 2017 Jun 26. Virus Res. 2018. PMID: 28655548 Free PMC article. Review.
-
Modulation of Dengue/Zika Virus Pathogenicity by Antibody-Dependent Enhancement and Strategies to Protect Against Enhancement in Zika Virus Infection.Front Immunol. 2018 Apr 23;9:597. doi: 10.3389/fimmu.2018.00597. eCollection 2018. Front Immunol. 2018. PMID: 29740424 Free PMC article. Review.
Cited by
-
A human iPSC-array-based GWAS identifies a virus susceptibility locus in the NDUFA4 gene and functional variants.Cell Stem Cell. 2022 Oct 6;29(10):1475-1490.e6. doi: 10.1016/j.stem.2022.09.008. Cell Stem Cell. 2022. PMID: 36206731 Free PMC article.
-
Memory B cell repertoire for recognition of evolving SARS-CoV-2 spike.bioRxiv [Preprint]. 2021 Mar 10:2021.03.10.434840. doi: 10.1101/2021.03.10.434840. bioRxiv. 2021. Update in: Cell. 2021 Sep 16;184(19):4969-4980.e15. doi: 10.1016/j.cell.2021.07.025. PMID: 33758863 Free PMC article. Updated. Preprint.
-
Development of a Potent and Protective Germline-Like Antibody Lineage Against Zika Virus in a Convalescent Human.Front Immunol. 2019 Oct 24;10:2424. doi: 10.3389/fimmu.2019.02424. eCollection 2019. Front Immunol. 2019. PMID: 31708914 Free PMC article.
-
A Gorilla Adenovirus-Based Vaccine against Zika Virus Induces Durable Immunity and Confers Protection in Pregnancy.Cell Rep. 2019 Sep 3;28(10):2634-2646.e4. doi: 10.1016/j.celrep.2019.08.005. Cell Rep. 2019. PMID: 31484074 Free PMC article.
-
Resurfaced ZIKV EDIII nanoparticle immunogens elicit neutralizing and protective responses in vivo.Cell Chem Biol. 2022 May 19;29(5):811-823.e7. doi: 10.1016/j.chembiol.2022.02.004. Epub 2022 Feb 28. Cell Chem Biol. 2022. PMID: 35231399 Free PMC article.
References
-
- Barba-Spaeth G, Dejnirattisai W, Rouvinski A, Vaney MC, Medits I, Sharma A, Simon-Loriere E, Sakuntabhai A, Cao-Lormeau VM, Haouz A, et al. Structural basis of potent Zika-dengue virus antibody cross-neutralization. Nature. 2016;536:48–53. - PubMed
-
- Barzon L, Pacenti M, Franchin E, Lavezzo E, Trevisan M, Sgarabotto D, Palu G. Infection dynamics in a traveller with persistent shedding of Zika virus RNA in semen for six months after returning from Haiti to Italy, January 2016. Euro surveillance : bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin 21 2016 - PMC - PubMed
MeSH terms
Substances
Grants and funding
- U01 AI088752/AI/NIAID NIH HHS/United States
- R01 AI037526/AI/NIAID NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- R21 AI112602/AI/NIAID NIH HHS/United States
- R25 TW009338/TW/FIC NIH HHS/United States
- R37 AI037526/AI/NIAID NIH HHS/United States
- R01 TW009504/TW/FIC NIH HHS/United States
- UM1 AI100663/AI/NIAID NIH HHS/United States
- T32 AI070084/AI/NIAID NIH HHS/United States
- U19 AI057229/AI/NIAID NIH HHS/United States
- UL1 TR001866/TR/NCATS NIH HHS/United States
- R01 AI124690/AI/NIAID NIH HHS/United States
- R01 AI121207/AI/NIAID NIH HHS/United States
- U19 AI111825/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
