

Molecular Mechanism of Substrate Processing by the Cdc48 ATPase Complex
- PMID: 28475898
- PMCID: PMC5751438
- DOI: 10.1016/j.cell.2017.04.020
Molecular Mechanism of Substrate Processing by the Cdc48 ATPase Complex
Abstract
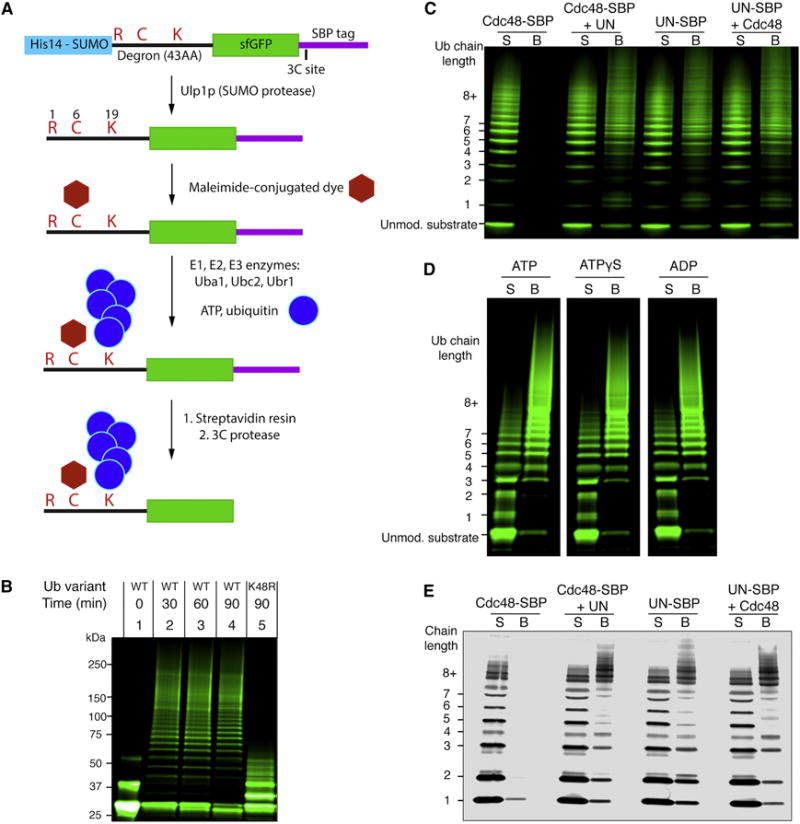
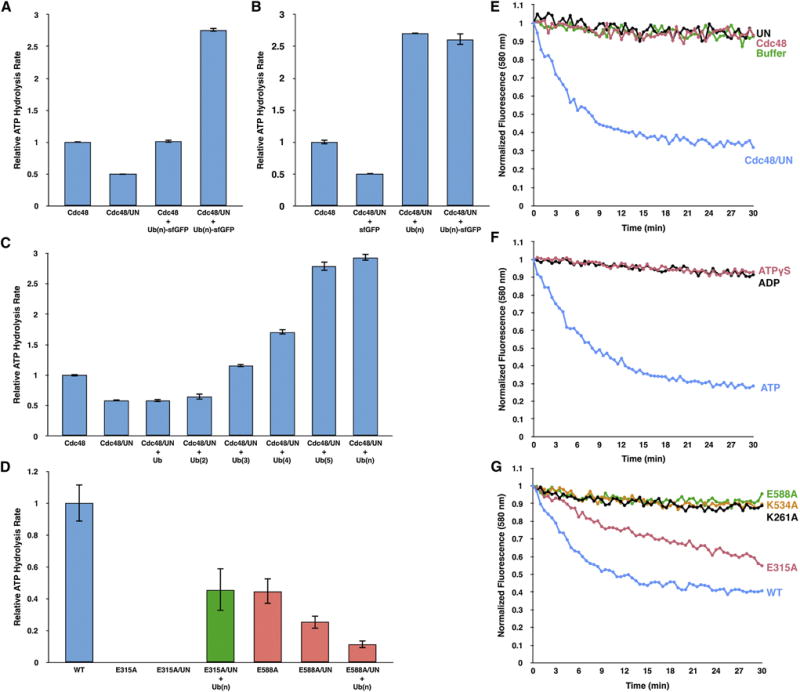
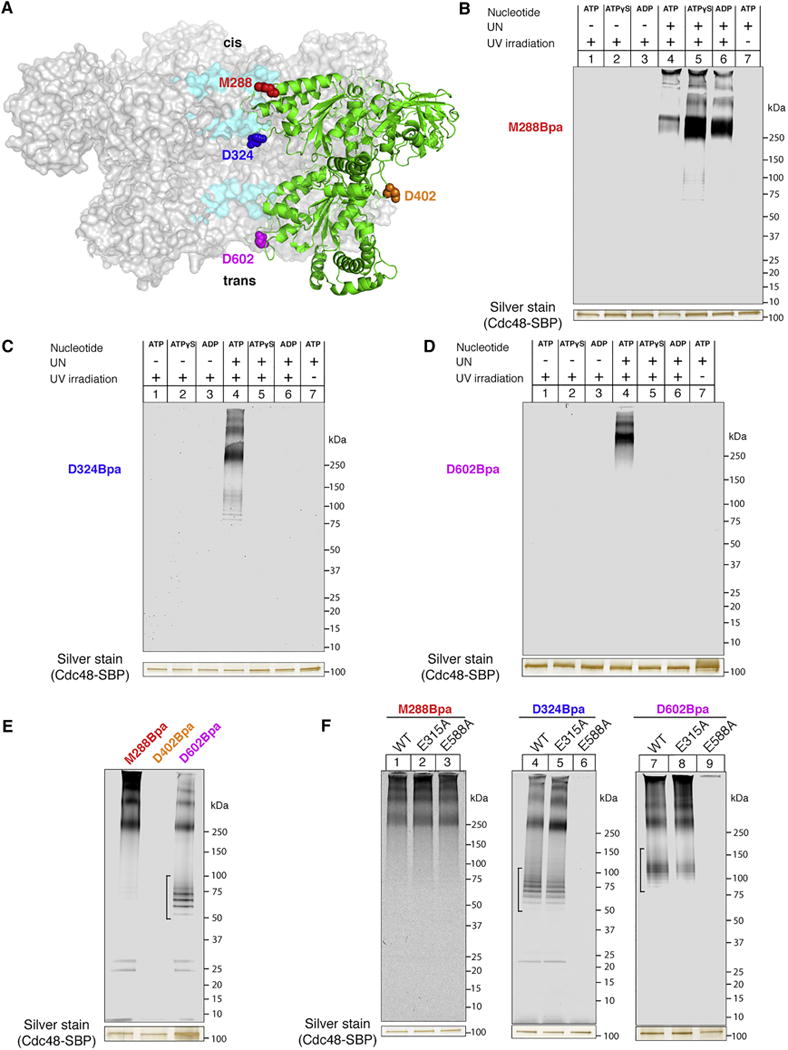
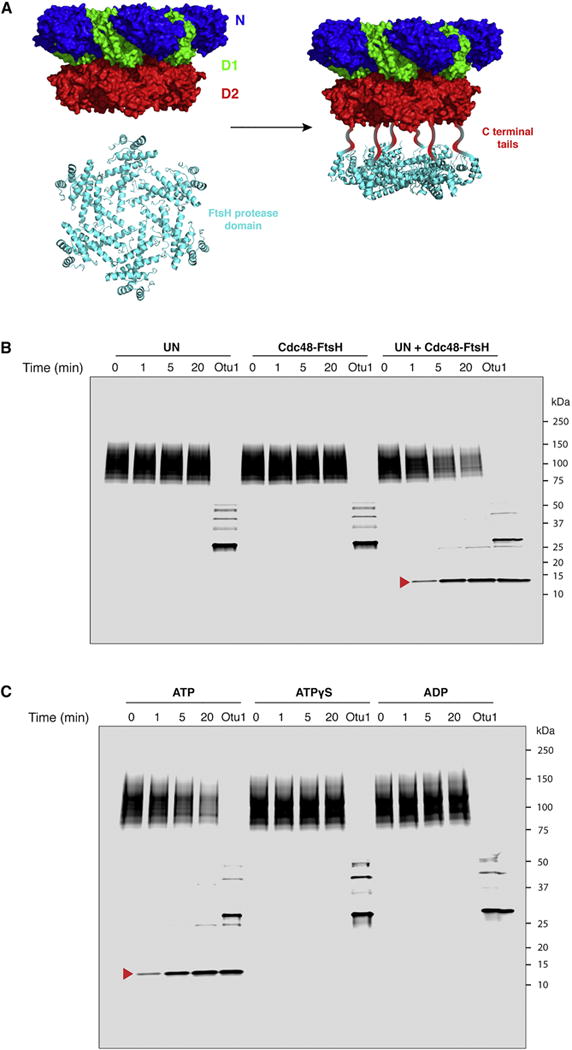
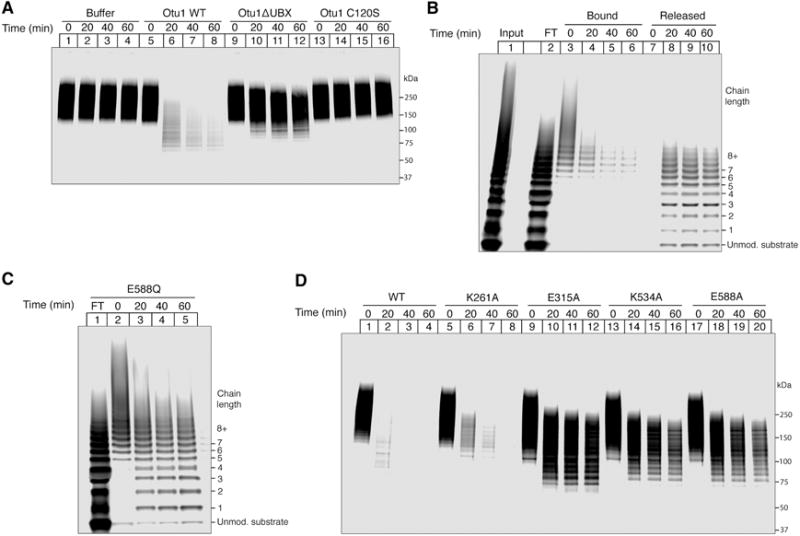
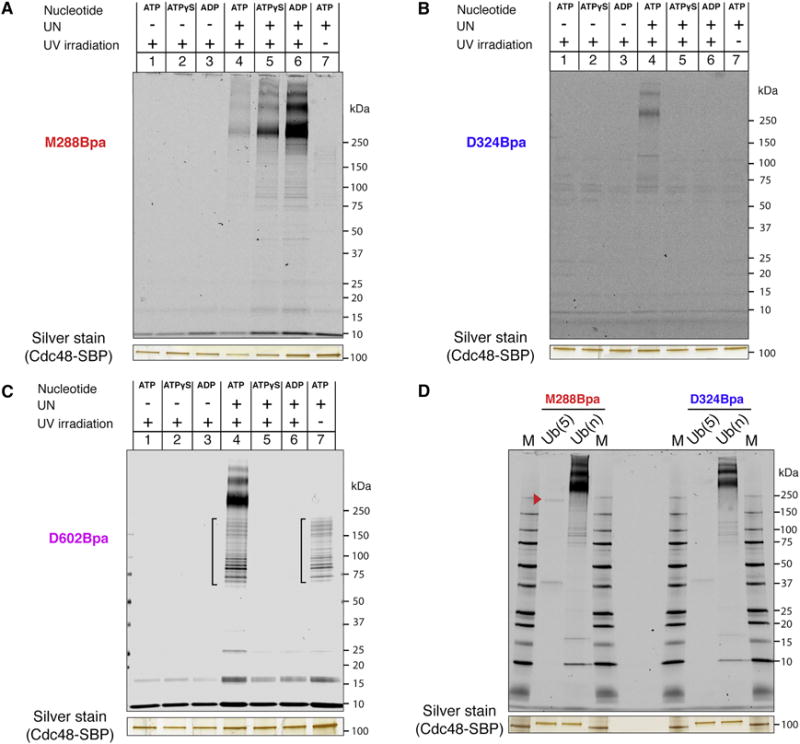
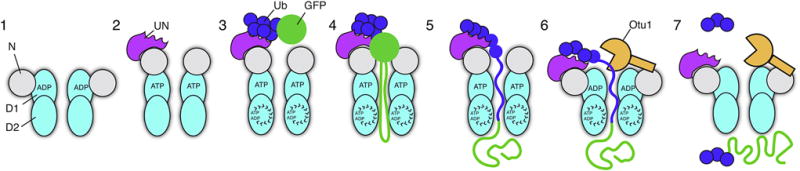
The Cdc48 ATPase and its cofactors Ufd1/Npl4 (UN) extract polyubiquitinated proteins from membranes or macromolecular complexes, but how they perform these functions is unclear. Cdc48 consists of an N-terminal domain that binds UN and two stacked hexameric ATPase rings (D1 and D2) surrounding a central pore. Here, we use purified components to elucidate how the Cdc48 complex processes substrates. After interaction of the polyubiquitin chain with UN, ATP hydrolysis by the D2 ring moves the polypeptide completely through the double ring, generating a pulling force on the substrate and causing its unfolding. ATP hydrolysis by the D1 ring is important for subsequent substrate release from the Cdc48 complex. This release requires cooperation of Cdc48 with a deubiquitinase, which trims polyubiquitin to an oligoubiquitin chain that is then also translocated through the pore. Together, these results lead to a new paradigm for the function of Cdc48 and its mammalian ortholog p97/VCP.
Keywords: AAA ATPase; ERAD; VCP; p97; proteasome; translocation; ubiuitination.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
