

Gating of Connexin Channels by transjunctional-voltage: Conformations and models of open and closed states
- PMID: 28476631
- PMCID: PMC5668201
- DOI: 10.1016/j.bbamem.2017.04.028
Gating of Connexin Channels by transjunctional-voltage: Conformations and models of open and closed states
Abstract
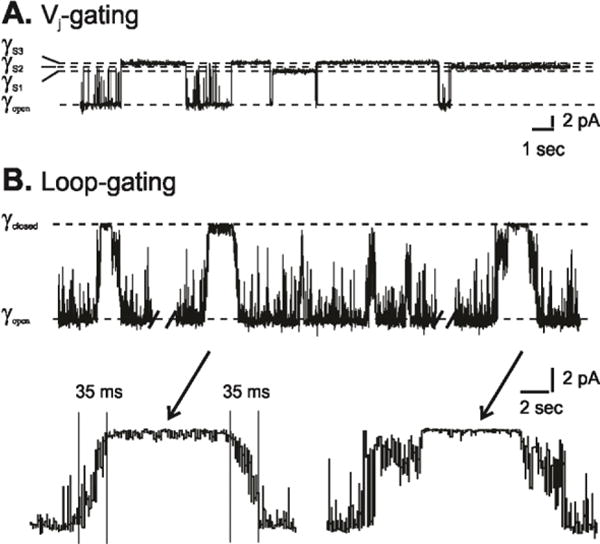
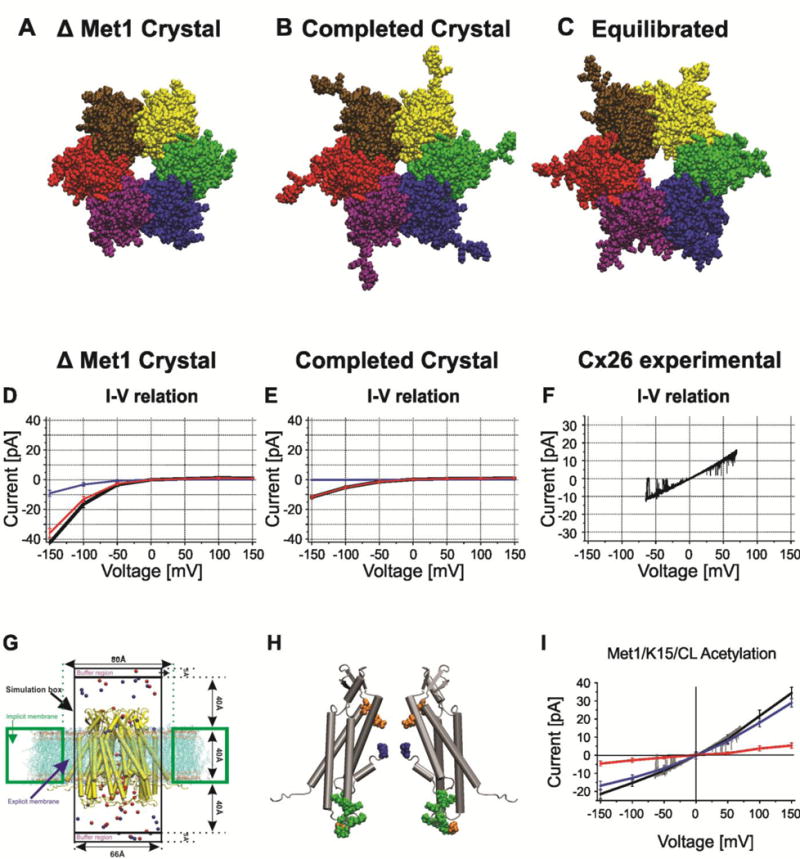
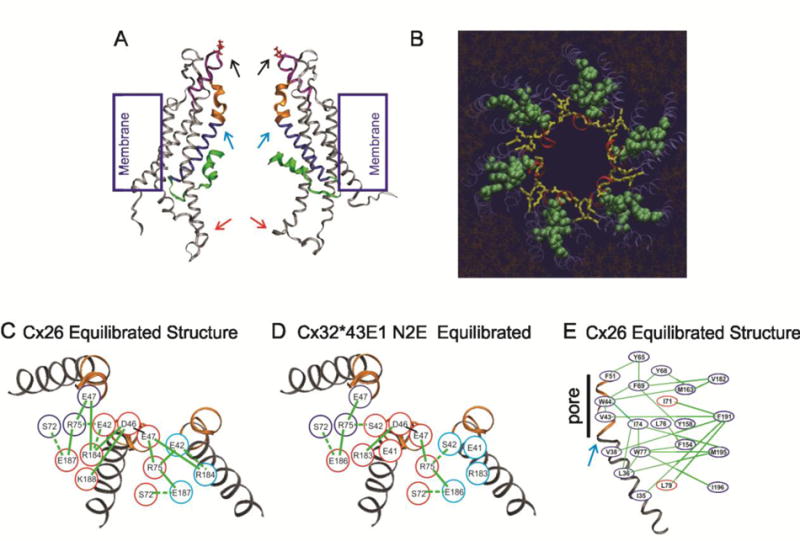
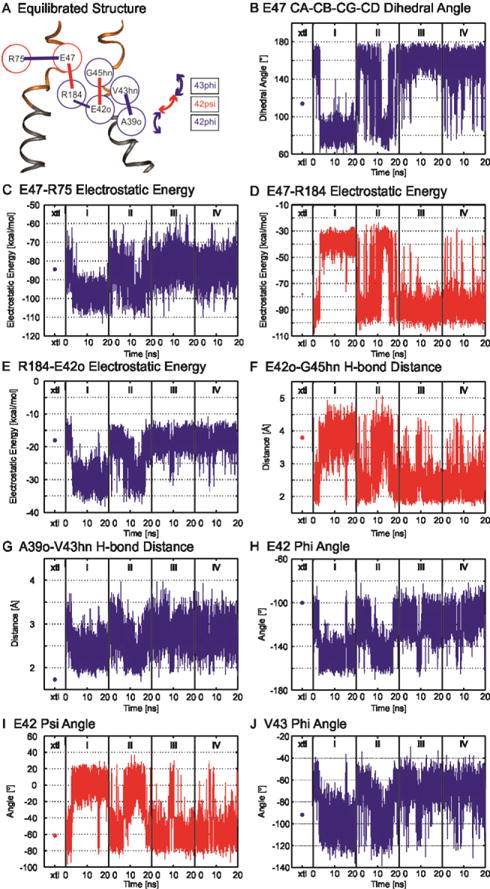
Voltage is an important physiologic regulator of channels formed by the connexin gene family. Connexins are unique among ion channels in that both plasma membrane inserted hemichannels (undocked hemichannels) and intercellular channels (aggregates of which form gap junctions) have important physiological roles. The hemichannel is the fundamental unit of gap junction voltage-gating. Each hemichannel displays two distinct voltage-gating mechanisms that are primarily sensitive to a voltage gradient formed along the length of the channel pore (the transjunctional voltage) rather than sensitivity to the absolute membrane potential (Vm or Vi-o). These transjunctional voltage dependent processes have been termed Vj- or fast-gating and loop- or slow-gating. Understanding the mechanism of voltage-gating, defined as the sequence of voltage-driven transitions that connect open and closed states, first and foremost requires atomic resolution models of the end states. Although ion channels formed by connexins were among the first to be characterized structurally by electron microscopy and x-ray diffraction in the early 1980's, subsequent progress has been slow. Much of the current understanding of the structure-function relations of connexin channels is based on two crystal structures of Cx26 gap junction channels. Refinement of crystal structure by all-atom molecular dynamics and incorporation of charge changing protein modifications has resulted in an atomic model of the open state that arguably corresponds to the physiologic open state. Obtaining validated atomic models of voltage-dependent closed states is more challenging, as there are currently no methods to solve protein structure while a stable voltage gradient is applied across the length of an oriented channel. It is widely believed that the best approach to solve the atomic structure of a voltage-gated closed ion channel is to apply different but complementary experimental and computational methods and to use the resulting information to derive a consensus atomic structure that is then subjected to rigorous validation. In this paper, we summarize our efforts to obtain and validate atomic models of the open and voltage-driven closed states of undocked connexin hemichannels. This article is part of a Special Issue entitled: Gap Junction Proteins edited by Jean Claude Herve.
Keywords: connexin; electrophysiology; homology models; metal bridging; molecular dynamics; voltage-gating.
Copyright © 2017 Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors have no conflict of interests.
Figures
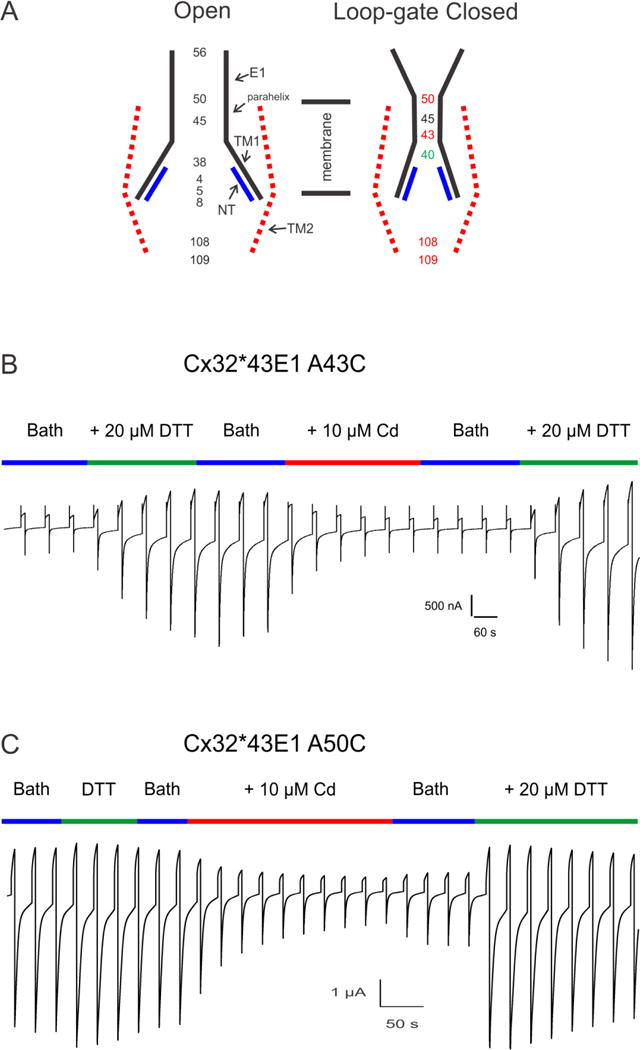
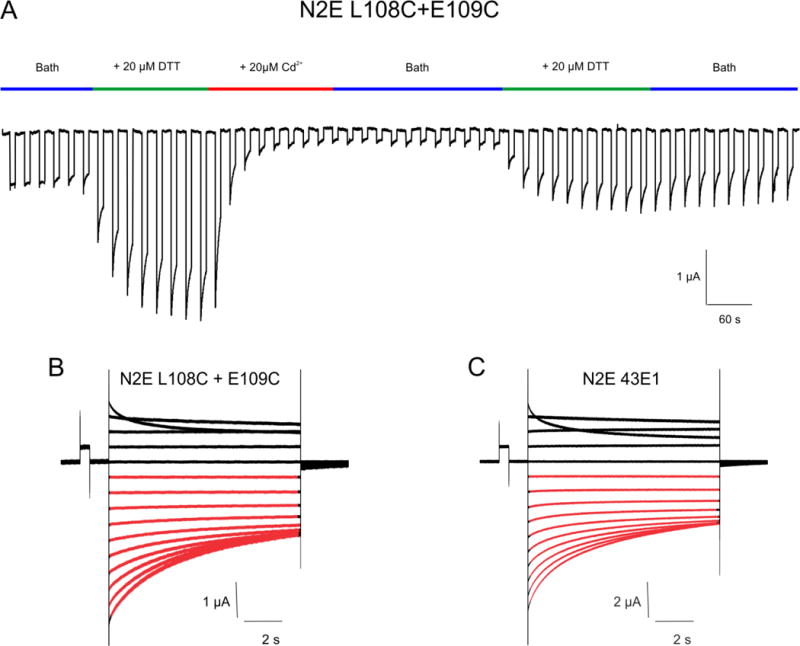
References
-
- Beyer EC, Berthoud VM. The Family of Connexin Genes. In: Harris AL, Locke D, editors. Connexins. Humana Press; 2009. pp. 3–26.
-
- Dewey MM, Barr L. Intercellular Connection between Smooth Muscle Cells: the Nexus. Science. 1962;137:670–672. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
