

Theta Oscillations during Active Sleep Synchronize the Developing Rubro-Hippocampal Sensorimotor Network
- PMID: 28479324
- PMCID: PMC5446093
- DOI: 10.1016/j.cub.2017.03.077
Theta Oscillations during Active Sleep Synchronize the Developing Rubro-Hippocampal Sensorimotor Network
Abstract
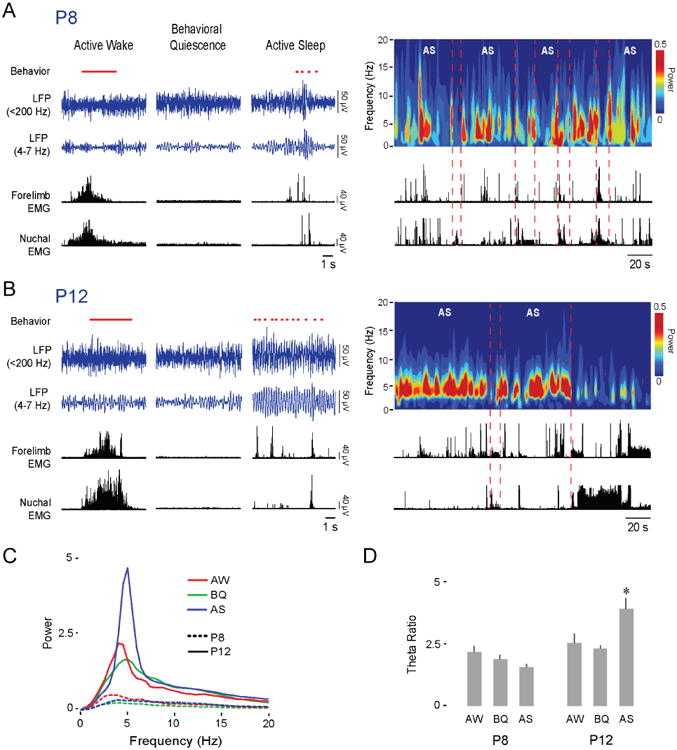
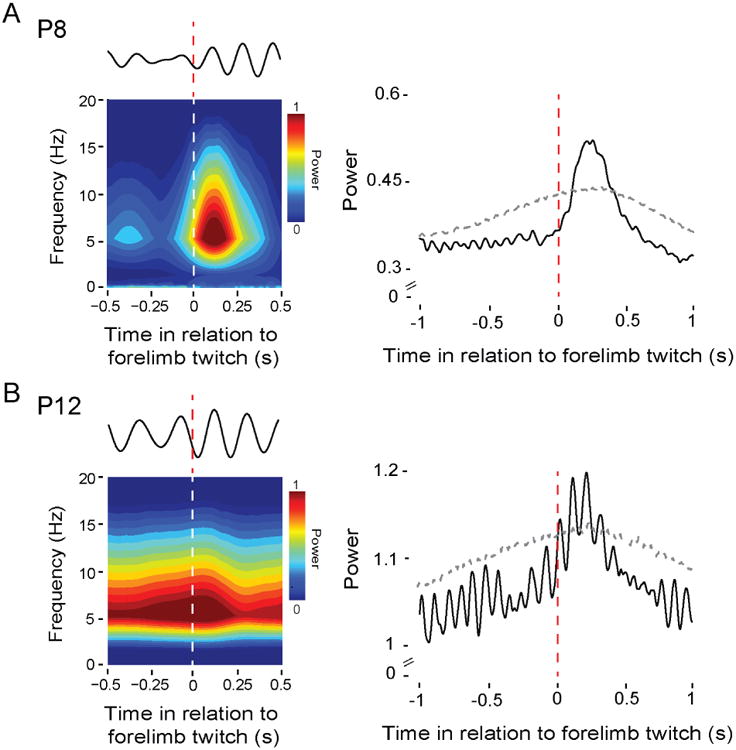
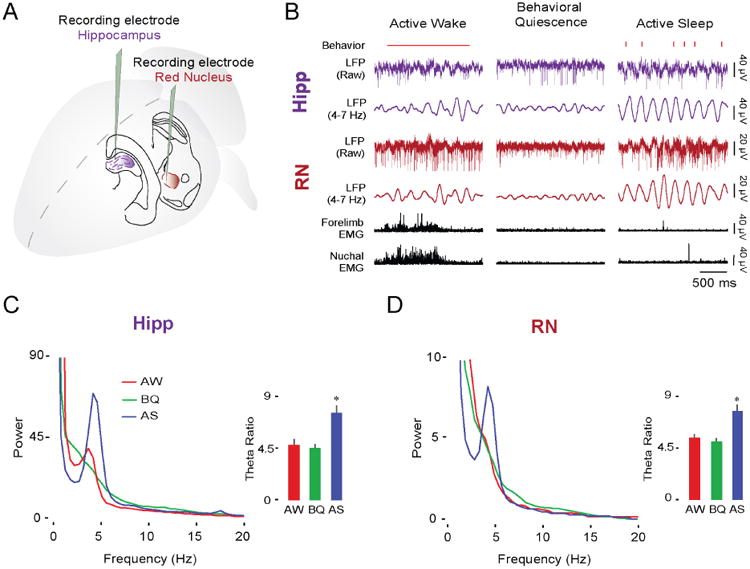
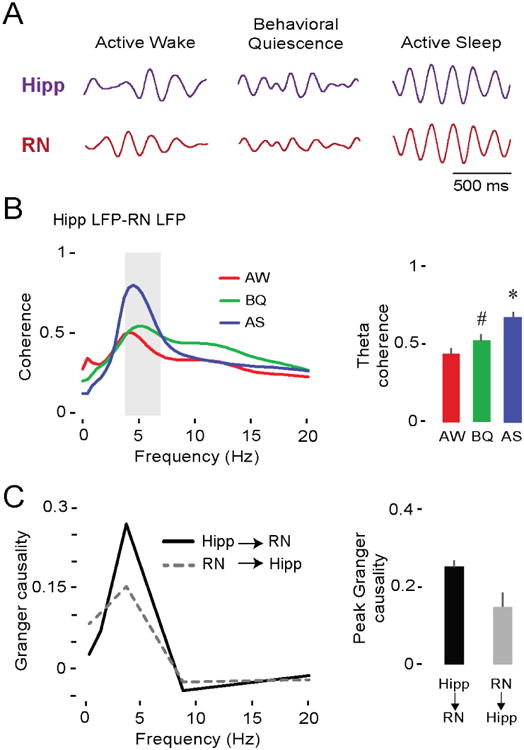
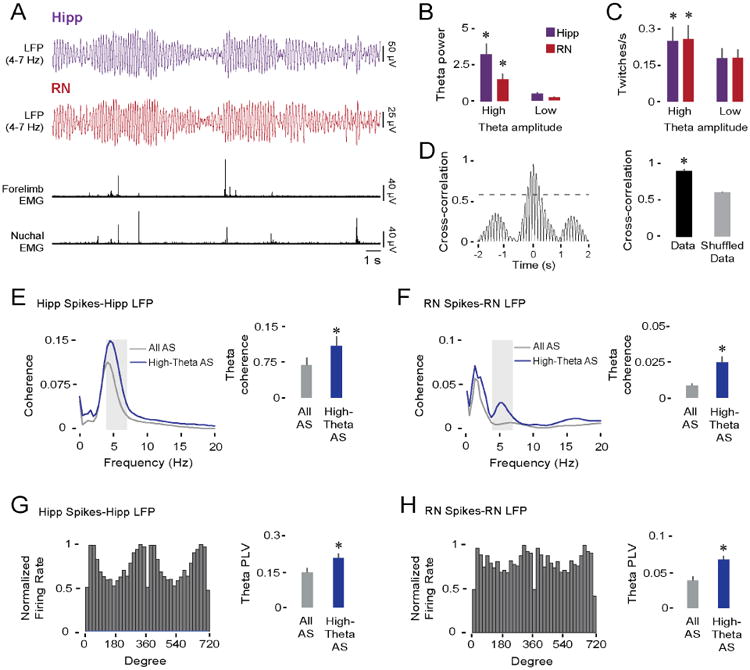
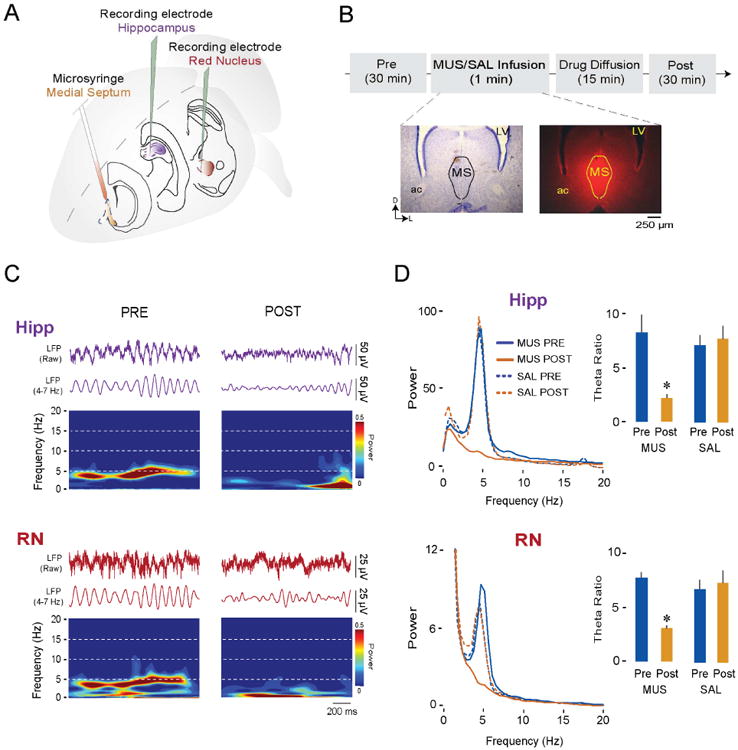
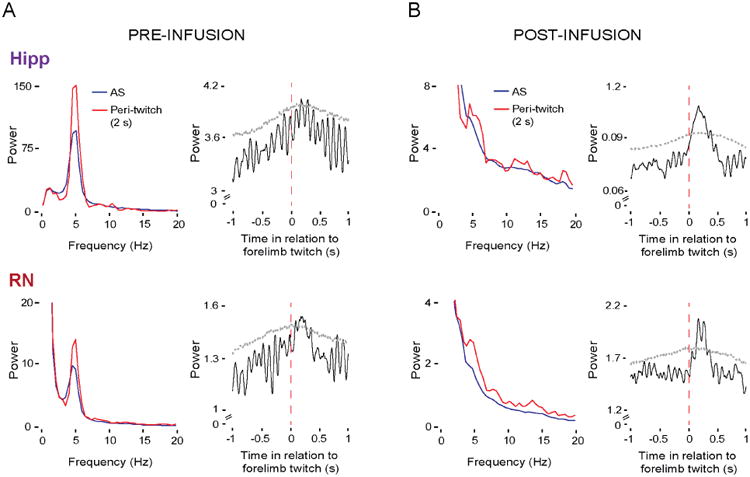
Neuronal oscillations comprise a fundamental mechanism by which distant neural structures establish and express functional connectivity. Long-range functional connectivity between the hippocampus and other forebrain structures is enabled by theta oscillations. Here, we show for the first time that the infant rat red nucleus (RN)-a brainstem sensorimotor structure-exhibits theta (4-7 Hz) oscillations restricted primarily to periods of active (REM) sleep. At postnatal day 8 (P8), theta is expressed as brief bursts immediately following myoclonic twitches; by P12, theta oscillations are expressed continuously across bouts of active sleep. Simultaneous recordings from the hippocampus and RN at P12 show that theta oscillations in both structures are coherent, co-modulated, and mutually interactive during active sleep. Critically, at P12, inactivation of the medial septum eliminates theta in both structures. The developmental emergence of theta-dependent functional coupling between the hippocampus and RN parallels that between the hippocampus and prefrontal cortex. Accordingly, disruptions in the early expression of theta could underlie the cognitive and sensorimotor deficits associated with neurodevelopmental disorders such as autism and schizophrenia.
Keywords: LFP; autism; connectivity; development; hippocampus; myoclonic twitching; neurodevelopmental disorder; red nucleus; sleep; theta rhythm.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Roffwarg HP, Muzio JN, Dement WC. Ontogenetic development of the human sleep-dream cycle. Science. 1966;152:604–619. - PubMed
-
- Jouvet-Mounier D, Astic L, Lacote D. Ontogenesis of the states of sleep in rat, cat, and guinea pig during the first postnatal month. Dev Psychobiol. 1970;2:216–239. - PubMed
-
- Blumberg MS, Seelke AMH. The form and function of infant sleep: From muscle to neocortex. In: Blumberg MS, Freeman JH, Robinson SR, editors. The Oxford Handbook of Developmental Behavioral Neuroscience. New York: Oxford University Press; 2010. pp. 391–423.
-
- Frank MG. Sleep and developmental plasticity: Not just for kids. Prog Brain Res. 2011;193:221–232. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
