

Plasticity, dynamics, and inhibition of emerging tetracycline resistance enzymes
- PMID: 28481346
- PMCID: PMC5478473
- DOI: 10.1038/nchembio.2376
Plasticity, dynamics, and inhibition of emerging tetracycline resistance enzymes
Abstract
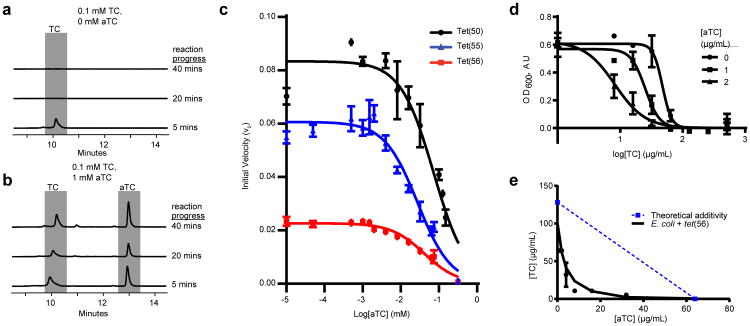
Although tetracyclines are an important class of antibiotics for use in agriculture and the clinic, their efficacy is threatened by increasing resistance. Resistance to tetracyclines can occur through efflux, ribosomal protection, or enzymatic inactivation. Surprisingly, tetracycline enzymatic inactivation has remained largely unexplored, despite providing the distinct advantage of antibiotic clearance. The tetracycline destructases are a recently discovered family of tetracycline-inactivating flavoenzymes from pathogens and soil metagenomes that have a high potential for broad dissemination. Here, we show that tetracycline destructases accommodate tetracycline-class antibiotics in diverse and novel orientations for catalysis, and antibiotic binding drives unprecedented structural dynamics facilitating tetracycline inactivation. We identify a key inhibitor binding mode that locks the flavin adenine dinucleotide cofactor in an inactive state, functionally rescuing tetracycline activity. Our results reveal the potential of a new tetracycline and tetracycline destructase inhibitor combination therapy strategy to overcome resistance by enzymatic inactivation and restore the use of an important class of antibiotics.
Figures
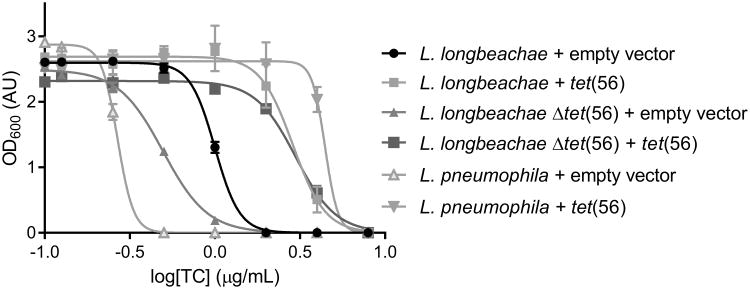

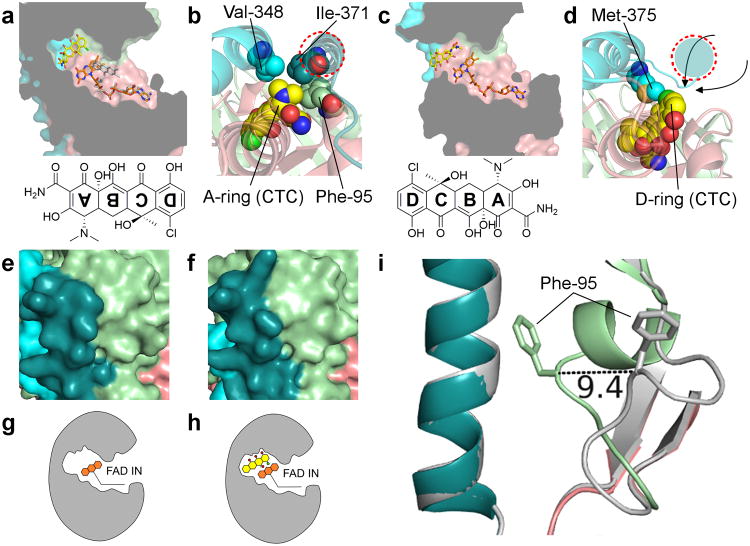
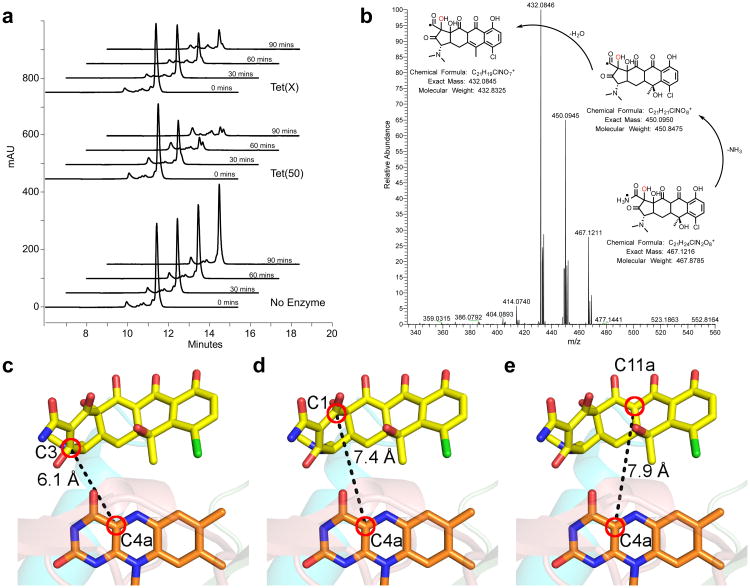
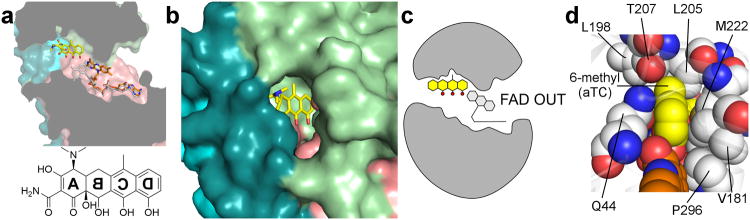
Comment in
-
Antibiotic resistance: Blocking tetracycline destruction.Nat Chem Biol. 2017 Jul;13(7):694-695. doi: 10.1038/nchembio.2396. Epub 2017 May 8. Nat Chem Biol. 2017. PMID: 28481345 No abstract available.
References
-
- Davies J. Inactivation of antibiotics and the dissemination of resistance genes. Science. 1994;264:375–382. - PubMed
online Methods References
-
- Berger KH, Isberg RR. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol Microbiol. 1993;7:7–19. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
