

Ectopic protein interactions within BRD4-chromatin complexes drive oncogenic megadomain formation in NUT midline carcinoma
- PMID: 28484033
- PMCID: PMC5448232
- DOI: 10.1073/pnas.1702086114
Ectopic protein interactions within BRD4-chromatin complexes drive oncogenic megadomain formation in NUT midline carcinoma
Abstract
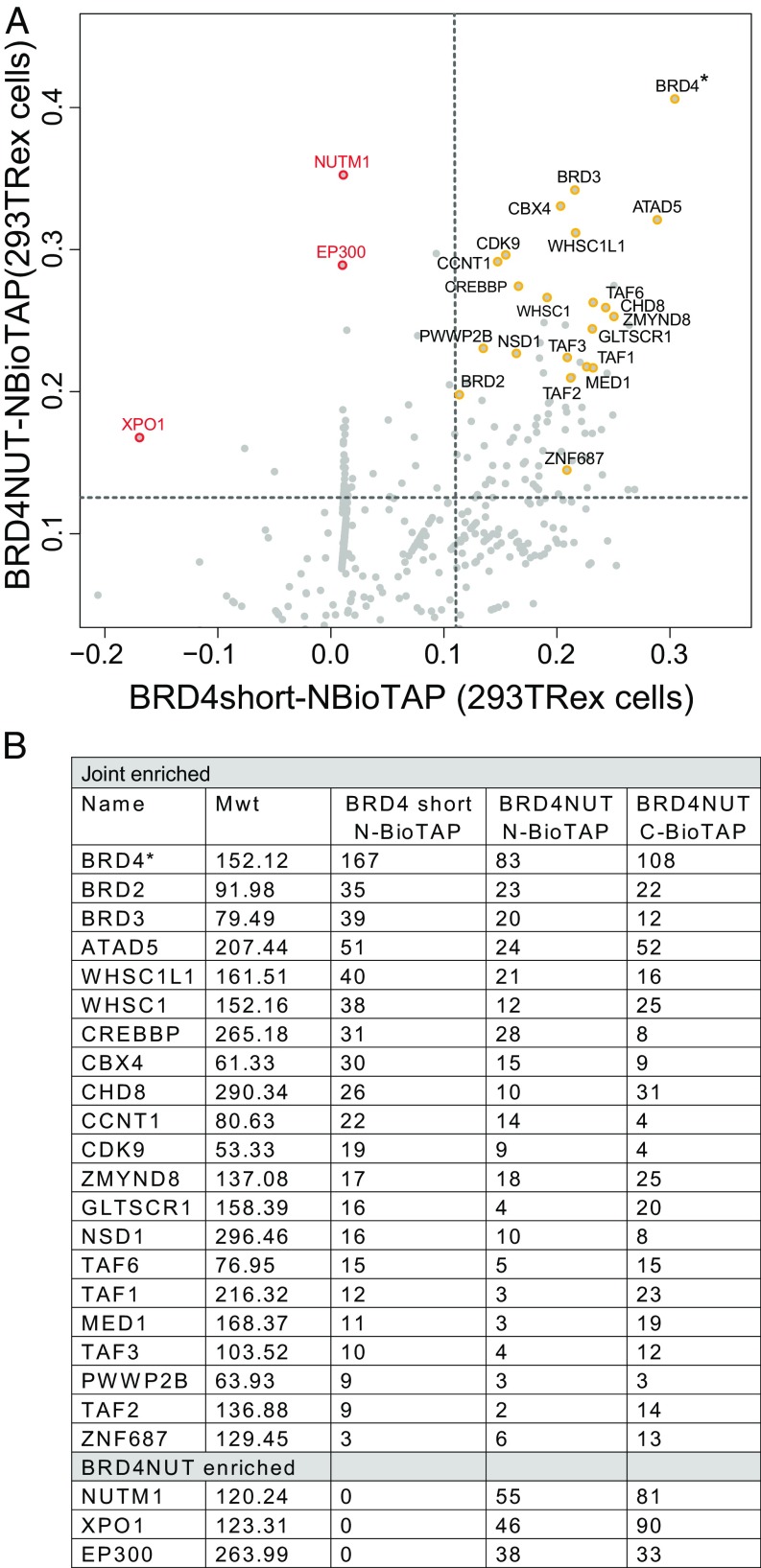
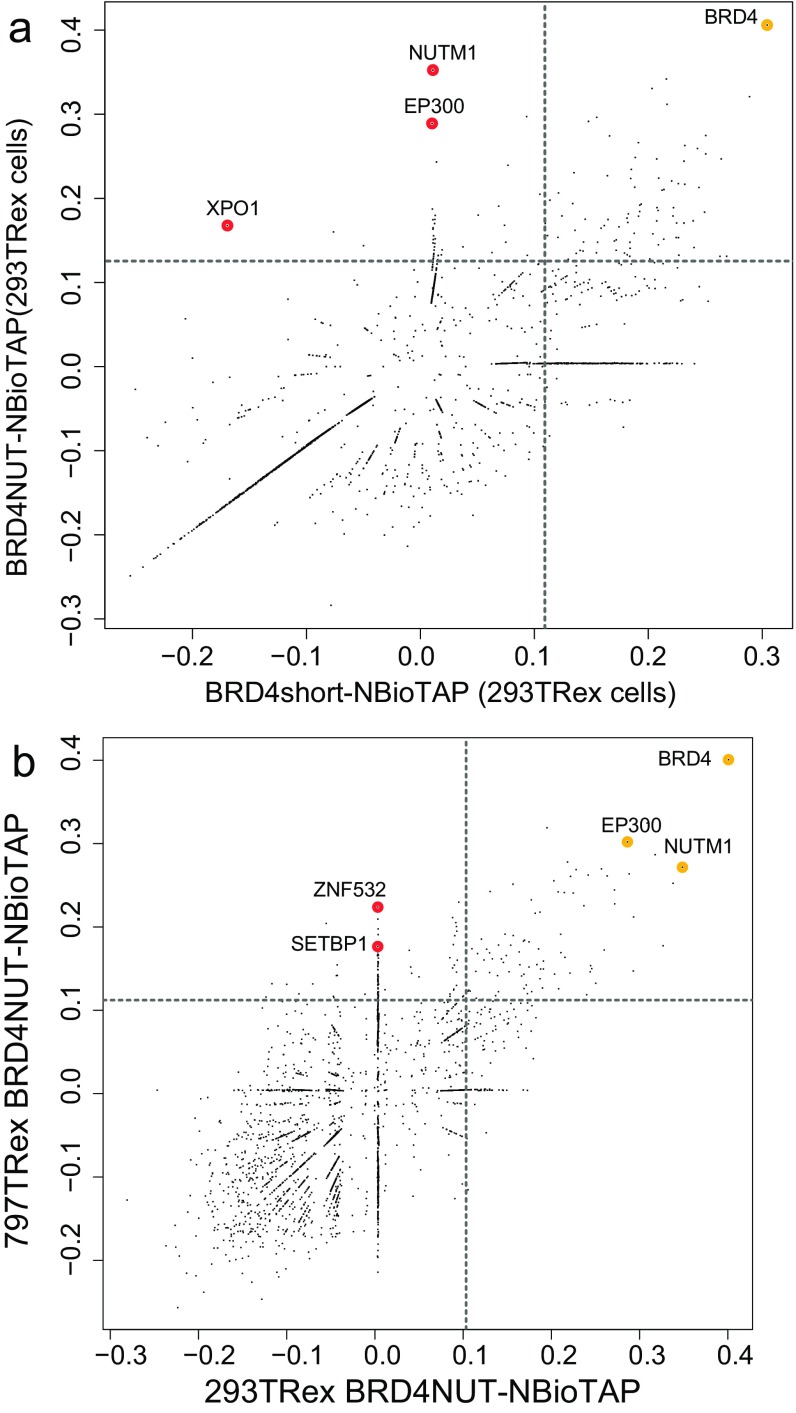
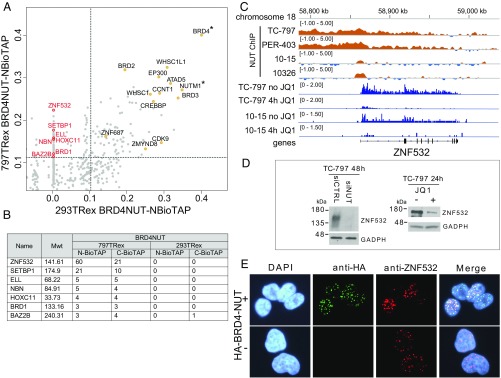
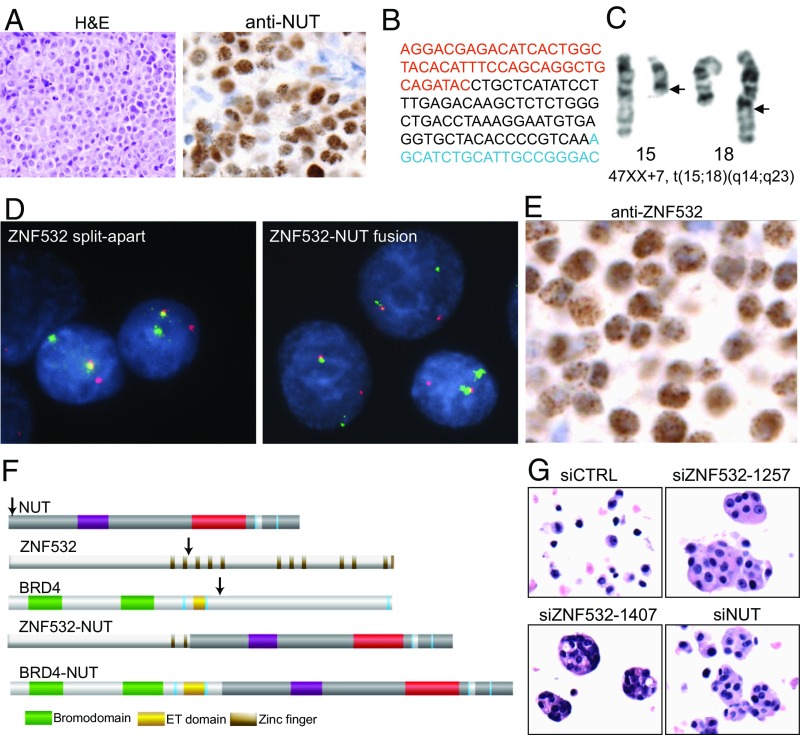
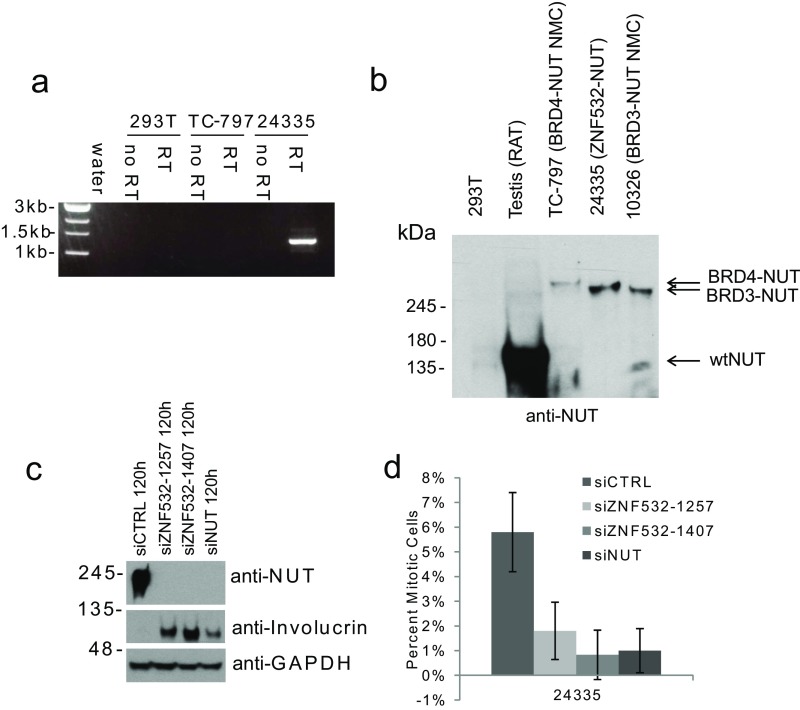
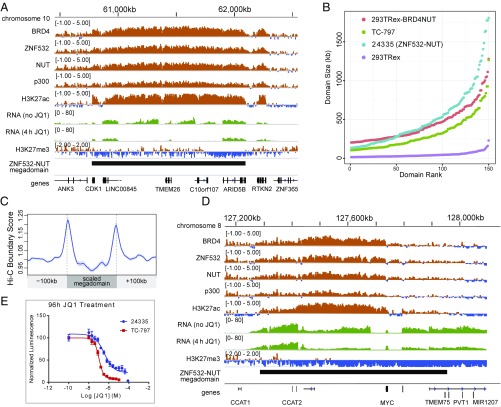
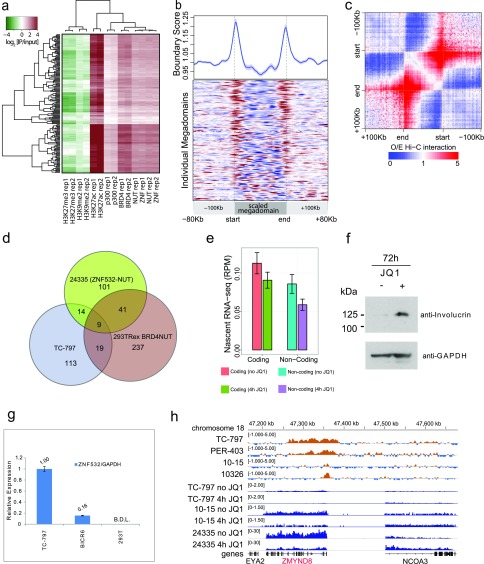
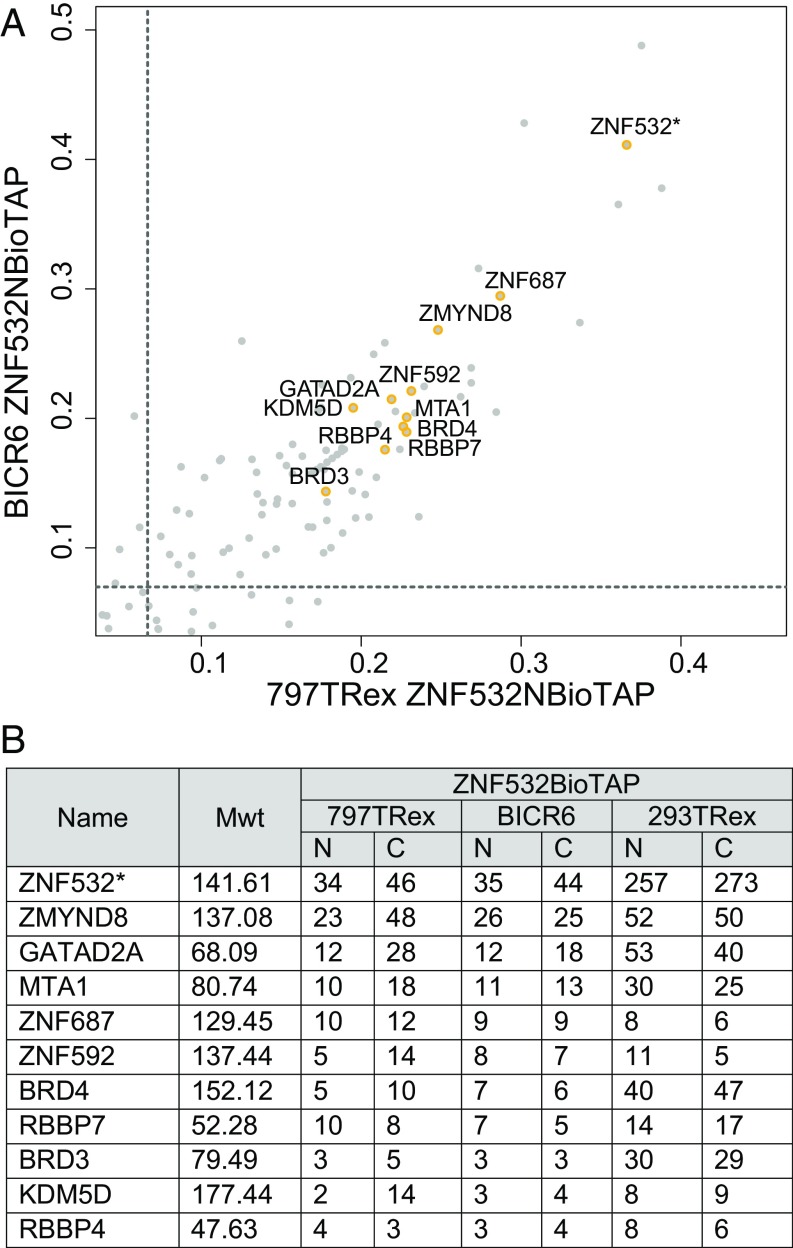
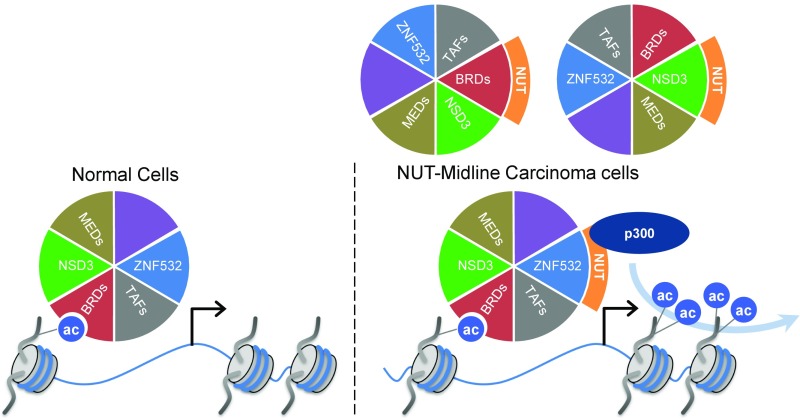
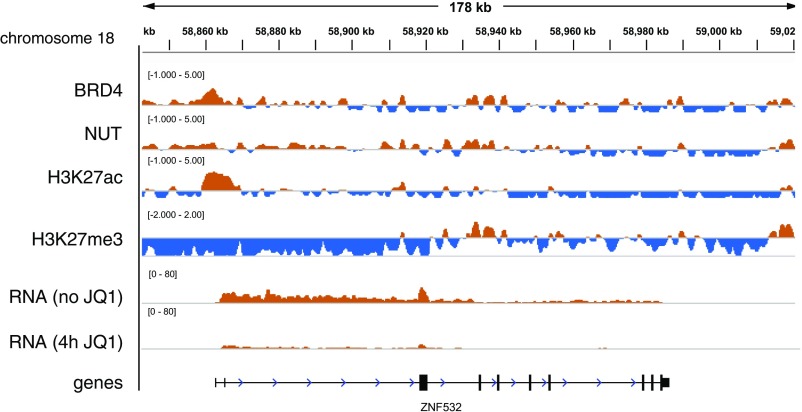
To investigate the mechanism that drives dramatic mistargeting of active chromatin in NUT midline carcinoma (NMC), we have identified protein interactions unique to the BRD4-NUT fusion oncoprotein compared with wild-type BRD4. Using cross-linking, affinity purification, and mass spectrometry, we identified the EP300 acetyltransferase as uniquely associated with BRD4 through the NUT fusion in both NMC and non-NMC cell types. We also discovered ZNF532 associated with BRD4-NUT in NMC patient cells but not detectable in 293T cells. EP300 and ZNF532 are both implicated in feed-forward regulatory loops leading to propagation of the oncogenic chromatin complex in BRD4-NUT patient cells. Adding key functional significance to our biochemical findings, we independently discovered a ZNF532-NUT translocation fusion in a newly diagnosed NMC patient. ChIP sequencing of the major players NUT, ZNF532, BRD4, EP300, and H3K27ac revealed the formation of ZNF532-NUT-associated hyperacetylated megadomains, distinctly localized but otherwise analogous to those found in BRD4-NUT patient cells. Our results support a model in which NMC is dependent on ectopic NUT-mediated interactions between EP300 and components of BRD4 regulatory complexes, leading to a cascade of misregulation.
Keywords: BRD4; BioTAP-XL; ZNF532–NUT; hyperacetylation; topological domains.
Conflict of interest statement
Conflict of interest statement: T.A.I. has a pending patent application relating to the OCMI-E medium.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
