

Cranial asymmetry arises later in the life history of the blind Mexican cavefish, Astyanax mexicanus
- PMID: 28486546
- PMCID: PMC5423691
- DOI: 10.1371/journal.pone.0177419
Cranial asymmetry arises later in the life history of the blind Mexican cavefish, Astyanax mexicanus
Abstract
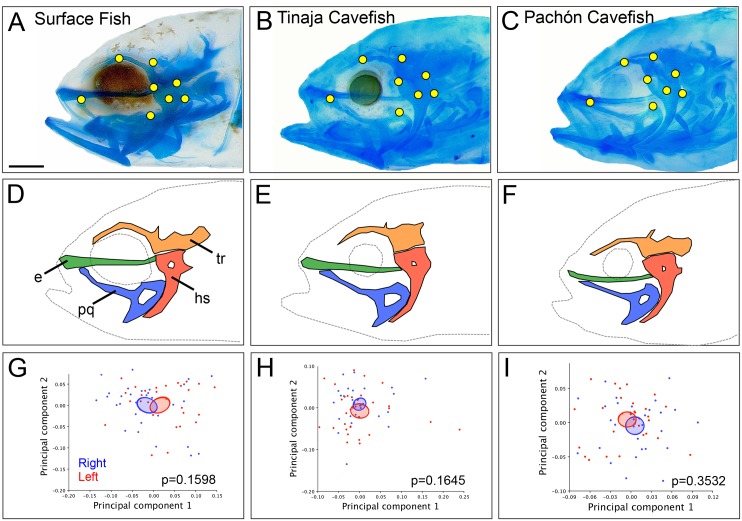
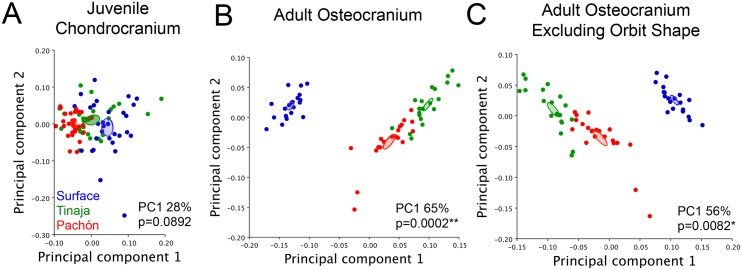
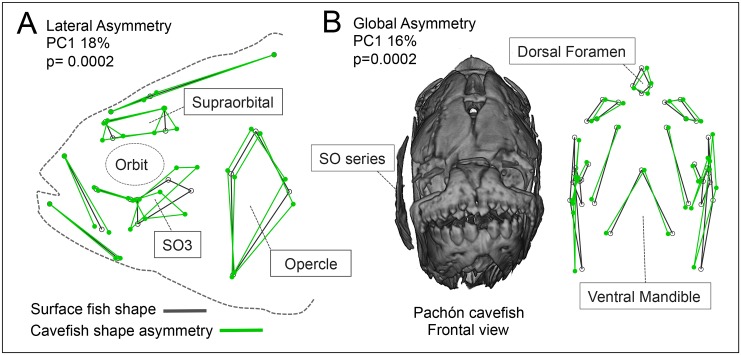
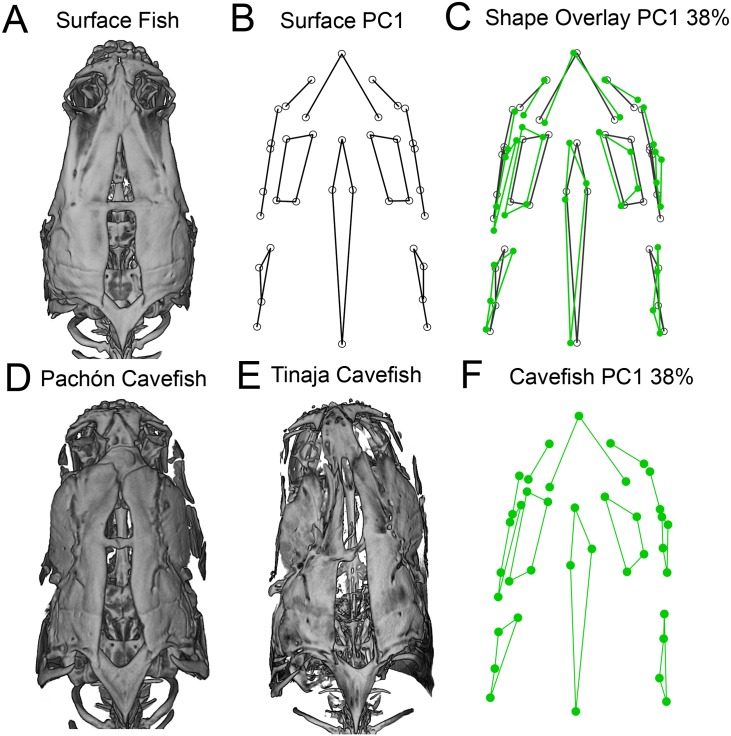
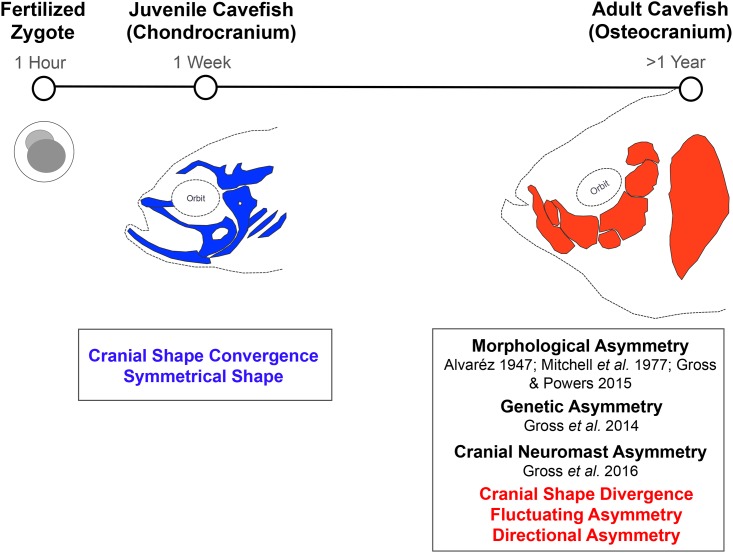
As a consequence of adaptation to the cave environment, the blind Mexican cavefish, Astyanax mexicanus, has evolved several cranial aberrations including changes to bone sizes, shapes and presence of numerous lateral asymmetries. Prior studies of cranial asymmetry in cavefish focused strictly on adult specimens. Thus, the extent to which these asymmetries emerge in adulthood, or earlier in the life history of cavefish, was unknown. We performed a geometric morphometric analysis of shape variation in the chondrocranium and osteocranium across life history in two distinct cavefish populations and surface-dwelling fish. The cartilaginous skull in juveniles was bilaterally symmetric and chondrocranial shape was conserved in all three populations. In contrast, bony skull shapes segregated into significantly distinct groups in adults. Cavefish demonstrated significant asymmetry for the bones surrounding the collapsed eye orbit, and the opercle bone posterior to the eye orbit. Interestingly, we discovered that cavefish also exhibit directional "bends" in skull shape, almost always biased to the left. In sum, this work reveals that asymmetric craniofacial aberrations emerge later in the cavefish life history. These abnormalities may mirror asymmetries in the lateral line sensory system, reflect a 'handedness' in cavefish swimming behavior, or evolve through neutral processes.
Conflict of interest statement
Figures
Similar articles
-
A Natural Animal Model System of Craniofacial Anomalies: The Blind Mexican Cavefish.Anat Rec (Hoboken). 2020 Jan;303(1):24-29. doi: 10.1002/ar.23998. Epub 2018 Nov 22. Anat Rec (Hoboken). 2020. PMID: 30365238 Free PMC article.
-
Parallel evolution of regressive and constructive craniofacial traits across distinct populations of Astyanax mexicanus cavefish.J Exp Zool B Mol Dev Evol. 2020 Nov;334(7-8):450-462. doi: 10.1002/jez.b.22932. Epub 2020 Feb 6. J Exp Zool B Mol Dev Evol. 2020. PMID: 32030873 Free PMC article.
-
Facial bone fragmentation in blind cavefish arises through two unusual ossification processes.Sci Rep. 2018 May 3;8(1):7015. doi: 10.1038/s41598-018-25107-2. Sci Rep. 2018. PMID: 29725043 Free PMC article.
-
Craniofacial skeleton of MEXICAN tetra (Astyanax mexicanus): As a bone disease model.Dev Dyn. 2019 Feb;248(2):153-161. doi: 10.1002/dvdy.4. Epub 2018 Dec 18. Dev Dyn. 2019. PMID: 30450697 Review.
-
Cavefish as a model system in evolutionary developmental biology.Dev Biol. 2001 Mar 1;231(1):1-12. doi: 10.1006/dbio.2000.0121. Dev Biol. 2001. PMID: 11180948 Review.
Cited by
-
A Natural Animal Model System of Craniofacial Anomalies: The Blind Mexican Cavefish.Anat Rec (Hoboken). 2020 Jan;303(1):24-29. doi: 10.1002/ar.23998. Epub 2018 Nov 22. Anat Rec (Hoboken). 2020. PMID: 30365238 Free PMC article.
-
An analysis of lateralized neural crest marker expression across development in the Mexican tetra, Astyanax mexicanus.Front Cell Dev Biol. 2023 Feb 15;11:1074616. doi: 10.3389/fcell.2023.1074616. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36875772 Free PMC article.
-
Variable Craniofacial Shape and Development among Multiple Cave-Adapted Populations of Astyanax mexicanus.Integr Org Biol. 2024 Aug 14;6(1):obae030. doi: 10.1093/iob/obae030. eCollection 2024. Integr Org Biol. 2024. PMID: 39234027 Free PMC article.
-
Maternal control of visceral asymmetry evolution in Astyanax cavefish.Sci Rep. 2021 May 13;11(1):10312. doi: 10.1038/s41598-021-89702-6. Sci Rep. 2021. PMID: 33986376 Free PMC article.
-
Genetic mapping of craniofacial traits in the Mexican tetra reveals loci associated with bite differences between cave and surface fish.BMC Ecol Evol. 2023 Aug 25;23(1):41. doi: 10.1186/s12862-023-02149-3. BMC Ecol Evol. 2023. PMID: 37626324 Free PMC article.
References
-
- Finnerty JR. The origins of axial patterning in the metazoa: how old is bilateral symmetry? Int J Dev Biol. 2003; 47:523–529. - PubMed
-
- Grammer K, Thornhill R.Human (Homo sapiens) facial attractiveness and sexual selection: the role of symmetry and averageness. J Comp Psy. 1994; 108:233–242. - PubMed
-
- Møller AP. Sexual selection in the barn swallow (Hirundo rustica). IV. Patterns of fluctuating asymmetry and selection against asymmetry. Evolution. 1994; 48:658–670. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
