

RNA-Dependent Cysteine Biosynthesis in Bacteria and Archaea
- PMID: 28487430
- PMCID: PMC5424206
- DOI: 10.1128/mBio.00561-17
RNA-Dependent Cysteine Biosynthesis in Bacteria and Archaea
Abstract
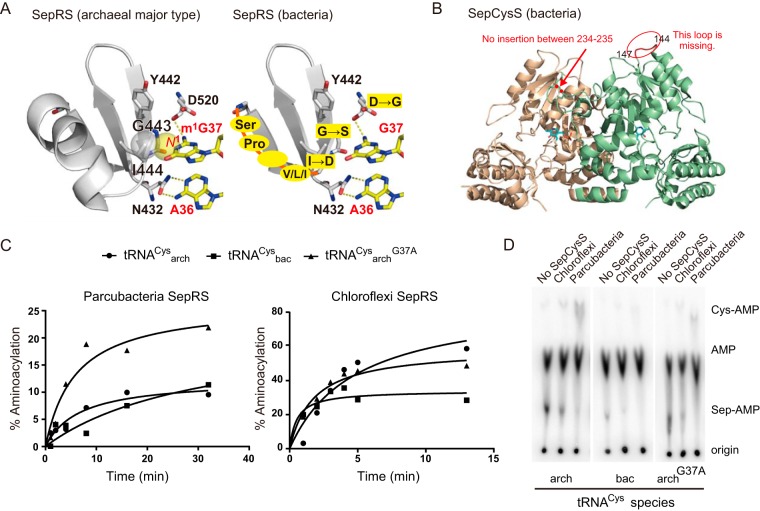
The diversity of the genetic code systems used by microbes on earth is yet to be elucidated. It is known that certain methanogenic archaea employ an alternative system for cysteine (Cys) biosynthesis and encoding; tRNACys is first acylated with phosphoserine (Sep) by O-phosphoseryl-tRNA synthetase (SepRS) and then converted to Cys-tRNACys by Sep-tRNA:Cys-tRNA synthase (SepCysS). In this study, we searched all genomic and metagenomic protein sequence data in the Integrated Microbial Genomes (IMG) system and at the NCBI to reveal new clades of SepRS and SepCysS proteins belonging to diverse archaea in the four major groups (DPANN, Euryarchaeota, TACK, and Asgard) and two groups of bacteria ("Candidatus Parcubacteria" and Chloroflexi). Bacterial SepRS and SepCysS charged bacterial tRNACys species with cysteine in vitro Homologs of SepCysE, a scaffold protein facilitating SepRS⋅SepCysS complex assembly in Euryarchaeota class I methanogens, are found in a few groups of TACK and Asgard archaea, whereas the C-terminally truncated homologs exist fused or genetically coupled with diverse SepCysS species. Investigation of the selenocysteine (Sec)- and pyrrolysine (Pyl)-utilizing traits in SepRS-utilizing archaea and bacteria revealed that the archaea carrying full-length SepCysE employ Sec and that SepRS is often found in Pyl-utilizing archaea and Chloroflexi bacteria. We discuss possible contributions of the SepRS-SepCysS system for sulfur assimilation, methanogenesis, and other metabolic processes requiring large amounts of iron-sulfur enzymes or Pyl-containing enzymes.IMPORTANCE Comprehensive analyses of all genomic and metagenomic protein sequence data in public databases revealed the distribution and evolution of an alternative cysteine-encoding system in diverse archaea and bacteria. The finding that the SepRS-SepCysS-SepCysE- and the selenocysteine-encoding systems are shared by the Euryarchaeota class I methanogens, the Crenarchaeota AK8/W8A-19 group, and an Asgard archaeon suggests that ancient archaea may have used both systems. In contrast, bacteria may have obtained the SepRS-SepCysS system from archaea. The SepRS-SepCysS system sometimes coexists with a pyrrolysine-encoding system in both archaea and bacteria. Our results provide additional bioinformatic evidence for the contribution of the SepRS-SepCysS system for sulfur assimilation and diverse metabolisms which require vast amounts of iron-sulfur enzymes and proteins. Among these biological activities, methanogenesis, methylamine metabolism, and organohalide respiration may have local and global effects on earth. Taken together, uncultured bacteria and archaea provide an expanded record of the evolution of the genetic code.
Keywords: biochemistry; bioinformatics; cysteine biosynthesis; genetic code; translation.
Copyright © 2017 Mukai et al.
Figures


Similar articles
-
RNA-dependent cysteine biosynthesis in archaea.Science. 2005 Mar 25;307(5717):1969-72. doi: 10.1126/science.1108329. Science. 2005. PMID: 15790858
-
Crystallographic analysis of a subcomplex of the transsulfursome with tRNA for Cys-tRNA(Cys) synthesis.Acta Crystallogr F Struct Biol Commun. 2016 Jul;72(Pt 7):569-72. doi: 10.1107/S2053230X16009559. Epub 2016 Jun 28. Acta Crystallogr F Struct Biol Commun. 2016. PMID: 27380375 Free PMC article.
-
Ancient translation factor is essential for tRNA-dependent cysteine biosynthesis in methanogenic archaea.Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):10520-5. doi: 10.1073/pnas.1411267111. Epub 2014 Jul 7. Proc Natl Acad Sci U S A. 2014. PMID: 25002468 Free PMC article.
-
From one amino acid to another: tRNA-dependent amino acid biosynthesis.Nucleic Acids Res. 2008 Apr;36(6):1813-25. doi: 10.1093/nar/gkn015. Epub 2008 Feb 5. Nucleic Acids Res. 2008. PMID: 18252769 Free PMC article. Review.
-
Biosynthesis of selenocysteine, the 21st amino acid in the genetic code, and a novel pathway for cysteine biosynthesis.Adv Nutr. 2011 Mar;2(2):122-8. doi: 10.3945/an.110.000265. Epub 2011 Mar 10. Adv Nutr. 2011. PMID: 22332041 Free PMC article. Review.
Cited by
-
Universal Codons with Enrichment from GC to AU Nucleotide Composition Reveal a Chronological Assignment from Early to Late Along with LUCA Formation.Life (Basel). 2020 Jun 5;10(6):81. doi: 10.3390/life10060081. Life (Basel). 2020. PMID: 32516985 Free PMC article.
-
New insights into the structure and function of an emerging drug target CysE.3 Biotech. 2021 Aug;11(8):373. doi: 10.1007/s13205-021-02891-9. Epub 2021 Jul 18. 3 Biotech. 2021. PMID: 34367865 Free PMC article. Review.
-
Methanomethylophilus alvus Mx1201 Provides Basis for Mutual Orthogonal Pyrrolysyl tRNA/Aminoacyl-tRNA Synthetase Pairs in Mammalian Cells.ACS Chem Biol. 2018 Nov 16;13(11):3087-3096. doi: 10.1021/acschembio.8b00571. Epub 2018 Oct 12. ACS Chem Biol. 2018. PMID: 30260624 Free PMC article.
-
Chemical Evolution of Life on Earth.Genes (Basel). 2025 Feb 13;16(2):220. doi: 10.3390/genes16020220. Genes (Basel). 2025. PMID: 40004549 Free PMC article. Review.
-
Possible Emergence of Sequence Specific RNA Aminoacylation via Peptide Intermediary to Initiate Darwinian Evolution and Code Through Origin of Life.Life (Basel). 2018 Oct 2;8(4):44. doi: 10.3390/life8040044. Life (Basel). 2018. PMID: 30279401 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
