

PD-1 regulates KLRG1+ group 2 innate lymphoid cells
- PMID: 28490441
- PMCID: PMC5461001
- DOI: 10.1084/jem.20161653
PD-1 regulates KLRG1+ group 2 innate lymphoid cells
Abstract
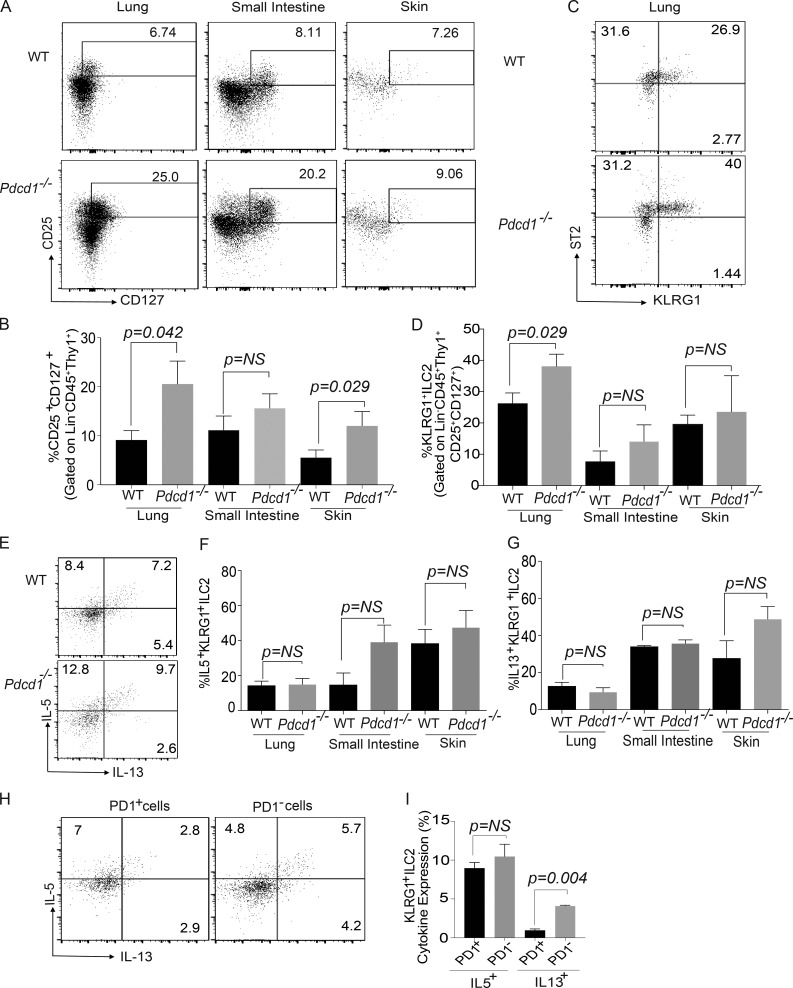
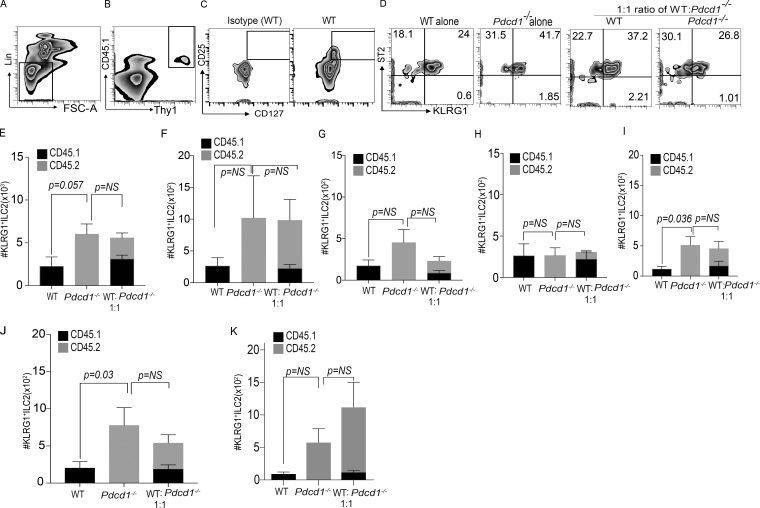
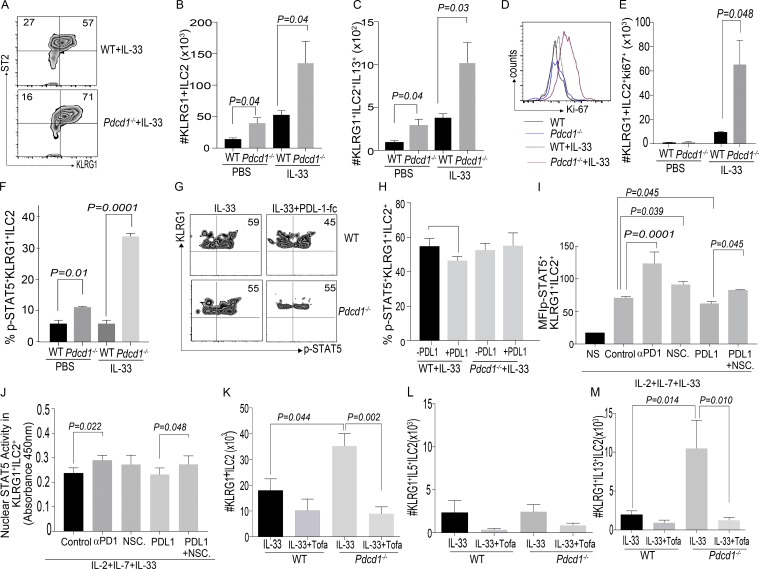
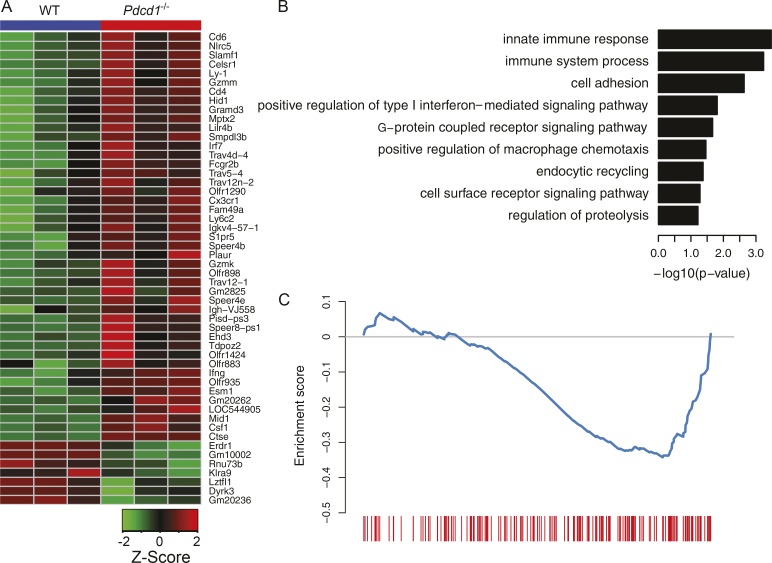
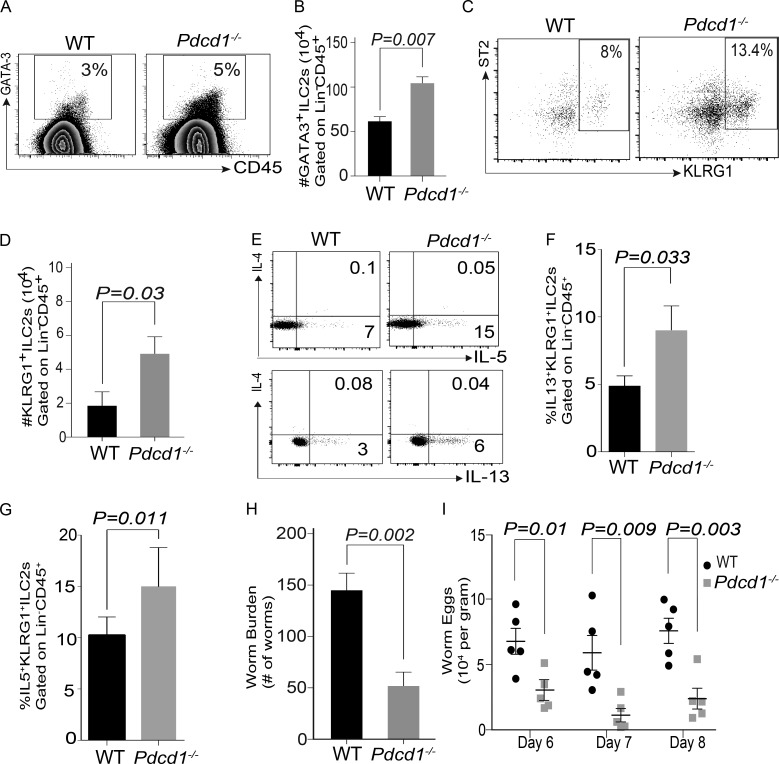
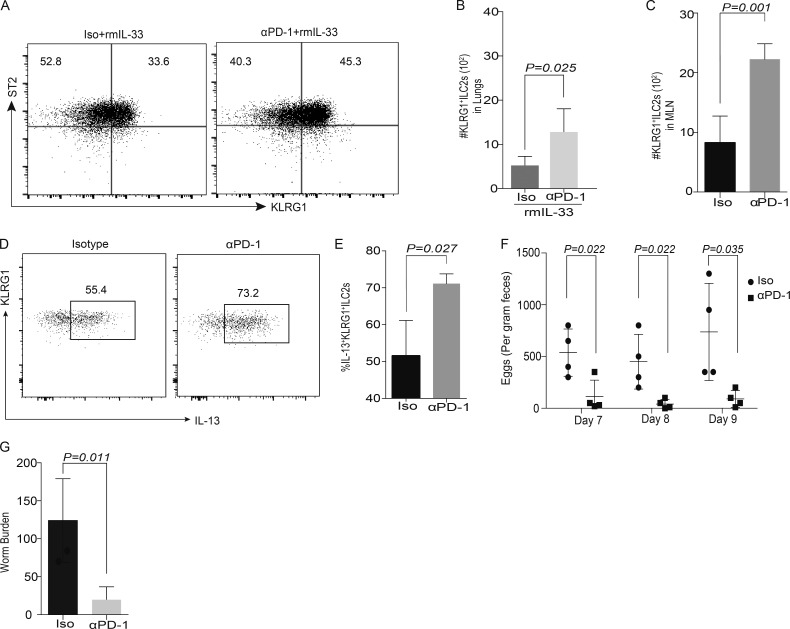
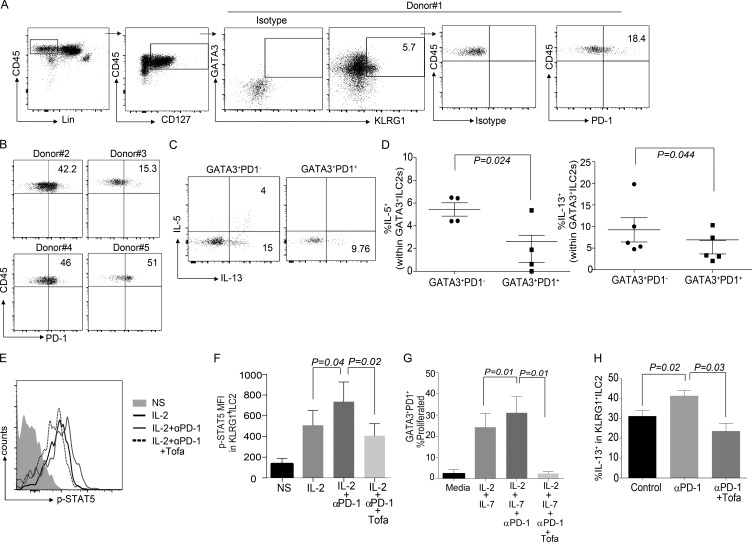
Group 2 innate lymphoid cells (ILC-2s) regulate immune responses to pathogens and maintain tissue homeostasis in response to cytokines. Positive regulation of ILC-2s through ICOS has been recently elucidated. We demonstrate here that PD-1 is an important negative regulator of KLRG1+ ILC-2 function in both mice and humans. Increase in KLRG1+ ILC-2 cell numbers was attributed to an intrinsic defect in PD-1 signaling, which resulted in enhanced STAT5 activation. During Nippostrongylus brasiliensis infection, a significant expansion of KLRG1+ ILC-2 subsets occurred in Pdcd1-/- mice and, upon adoptive transfer, Pdcd1-/- KLRG1+ ILC-2s significantly reduced worm burden. Furthermore, blocking PD-1 with an antibody increased KLRG1+ ILC-2 cell number and reduced disease burden. Therefore, PD-1 is required for maintaining the number, and hence function, of KLRG1+ ILC-2s.
© 2017 Taylor et al.
Figures
Comment in
-
Immune checkpoints on innate lymphoid cells.J Exp Med. 2017 Jun 5;214(6):1561-1563. doi: 10.1084/jem.20170763. Epub 2017 May 17. J Exp Med. 2017. PMID: 28515074 Free PMC article.
References
-
- Amarnath S., Costanzo C.M., Mariotti J., Ullman J.L., Telford W.G., Kapoor V., Riley J.L., Levine B.L., June C.H., Fong T., et al. 2010. Regulatory T cells and human myeloid dendritic cells promote tolerance via programmed death ligand-1. PLoS Biol. 8:e1000302 10.1371/journal.pbio.1000302 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
