

Entropic forces drive self-organization and membrane fusion by SNARE proteins
- PMID: 28490503
- PMCID: PMC5448213
- DOI: 10.1073/pnas.1611506114
Entropic forces drive self-organization and membrane fusion by SNARE proteins
Abstract
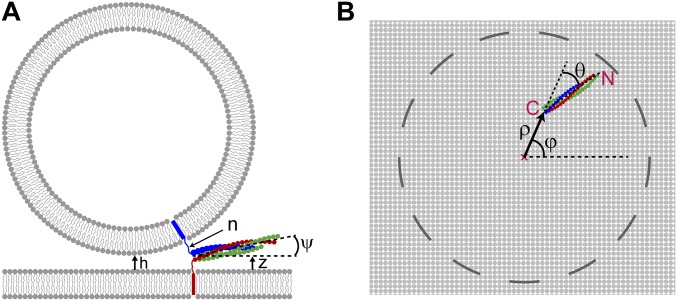
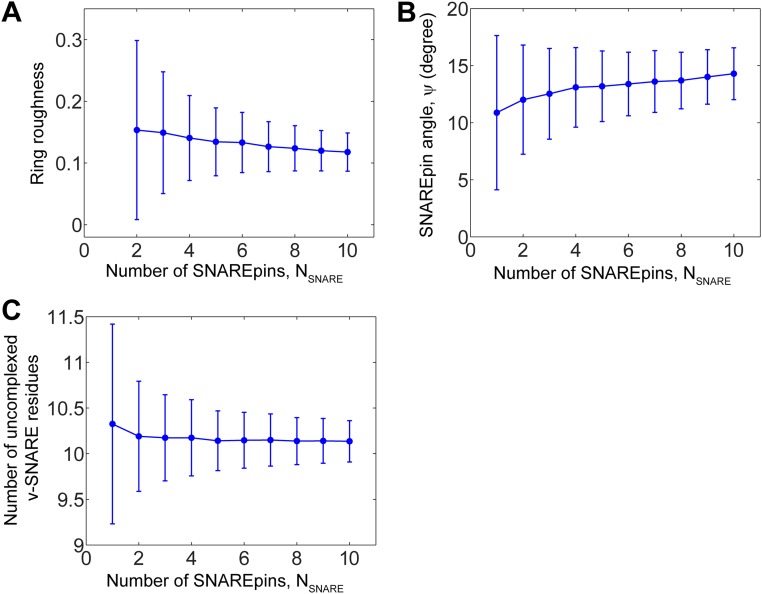
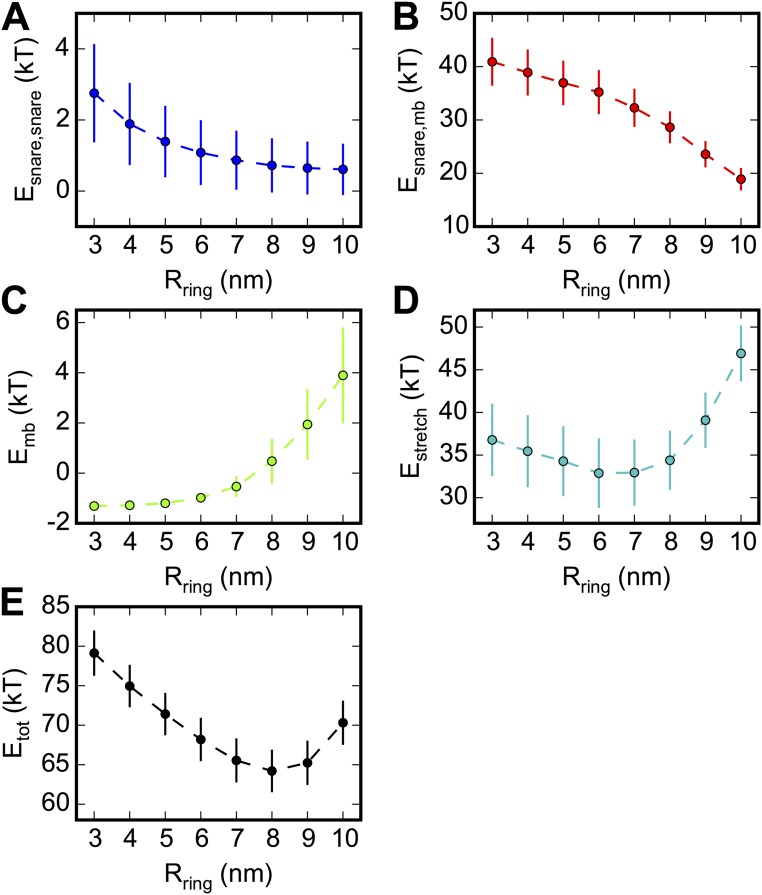
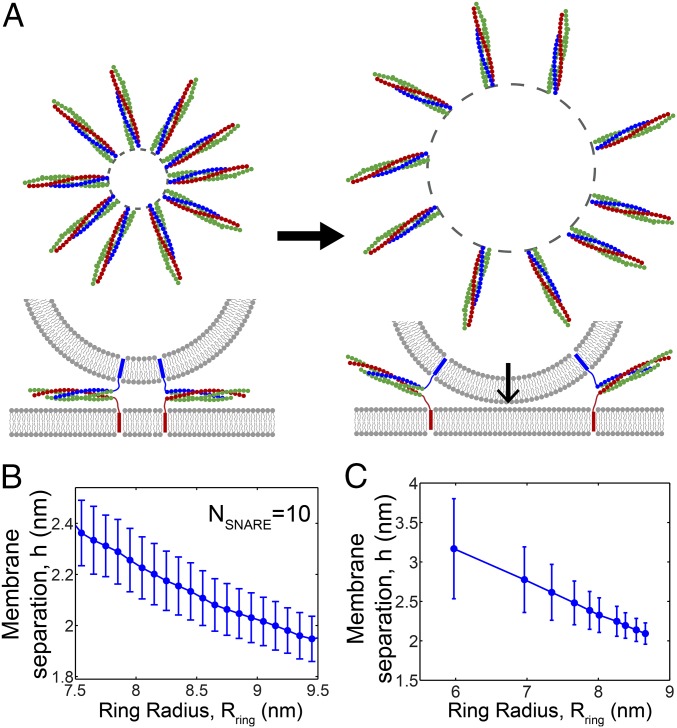
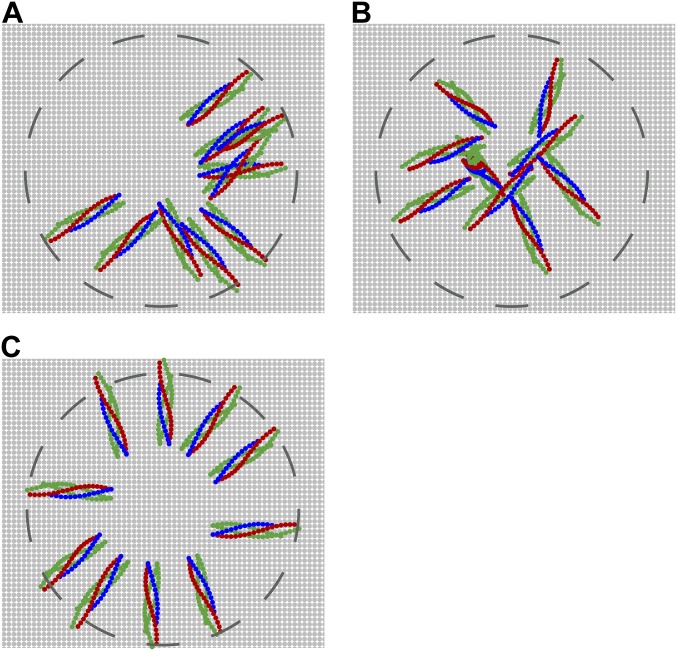
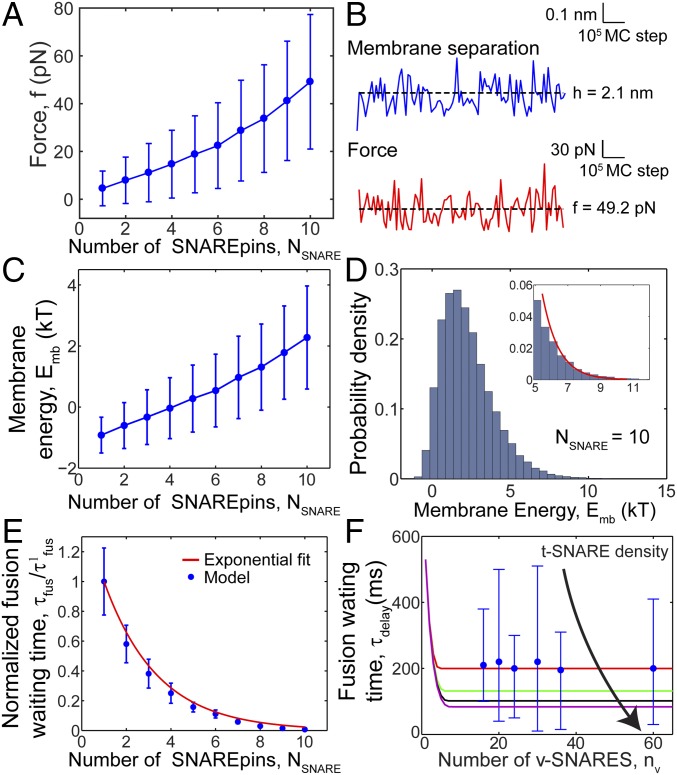
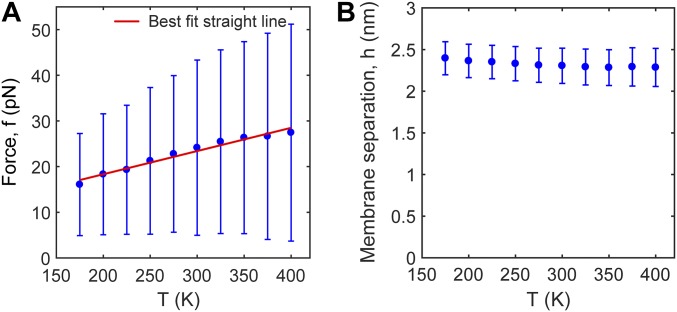
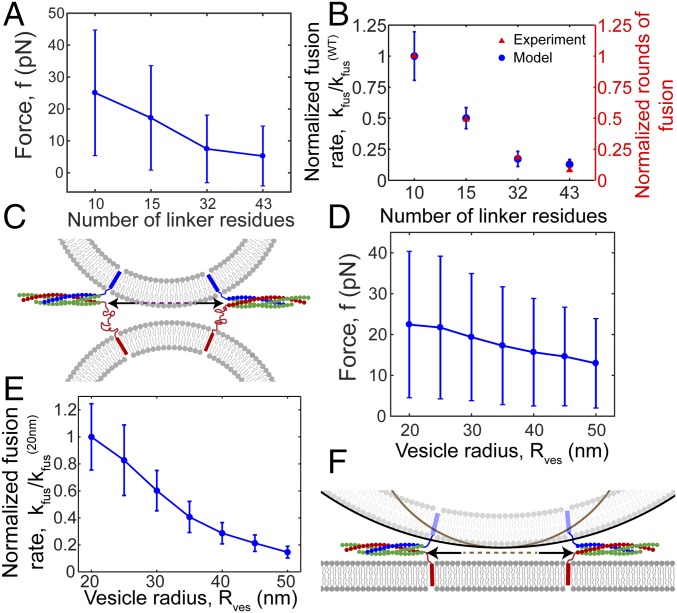
SNARE proteins are the core of the cell's fusion machinery and mediate virtually all known intracellular membrane fusion reactions on which exocytosis and trafficking depend. Fusion is catalyzed when vesicle-associated v-SNAREs form trans-SNARE complexes ("SNAREpins") with target membrane-associated t-SNAREs, a zippering-like process releasing ∼65 kT per SNAREpin. Fusion requires several SNAREpins, but how they cooperate is unknown and reports of the number required vary widely. To capture the collective behavior on the long timescales of fusion, we developed a highly coarse-grained model that retains key biophysical SNARE properties such as the zippering energy landscape and the surface charge distribution. In simulations the ∼65-kT zippering energy was almost entirely dissipated, with fully assembled SNARE motifs but uncomplexed linker domains. The SNAREpins self-organized into a circular cluster at the fusion site, driven by entropic forces that originate in steric-electrostatic interactions among SNAREpins and membranes. Cooperative entropic forces expanded the cluster and pulled the membranes together at the center point with high force. We find that there is no critical number of SNAREs required for fusion, but instead the fusion rate increases rapidly with the number of SNAREpins due to increasing entropic forces. We hypothesize that this principle finds physiological use to boost fusion rates to meet the demanding timescales of neurotransmission, exploiting the large number of v-SNAREs available in synaptic vesicles. Once in an unfettered cluster, we estimate ≥15 SNAREpins are required for fusion within the ∼1-ms timescale of neurotransmitter release.
Keywords: SNARE; entropic force; exocytosis; membrane fusion; neurotransmitter release.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
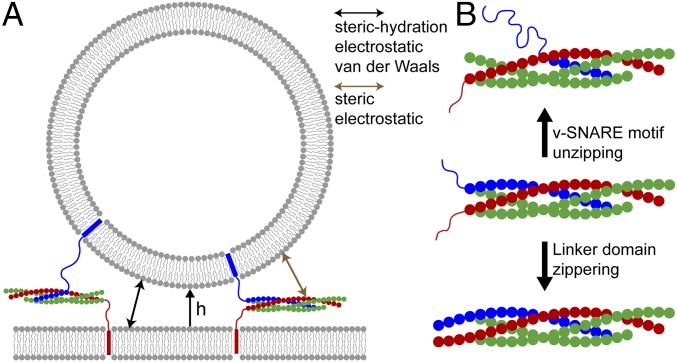
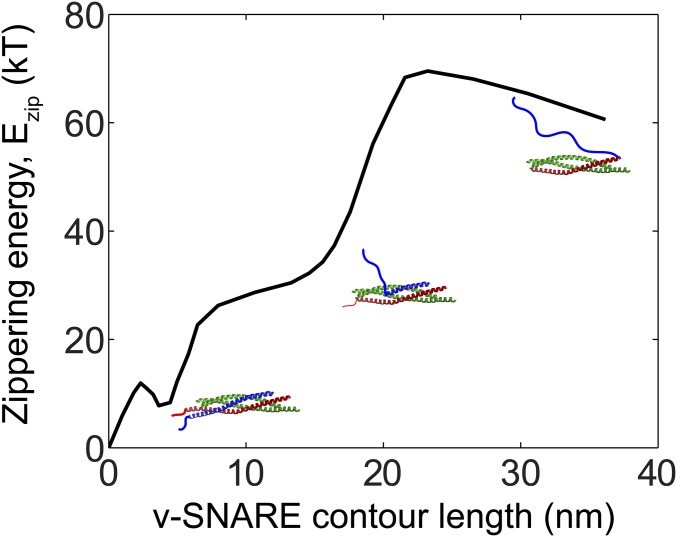
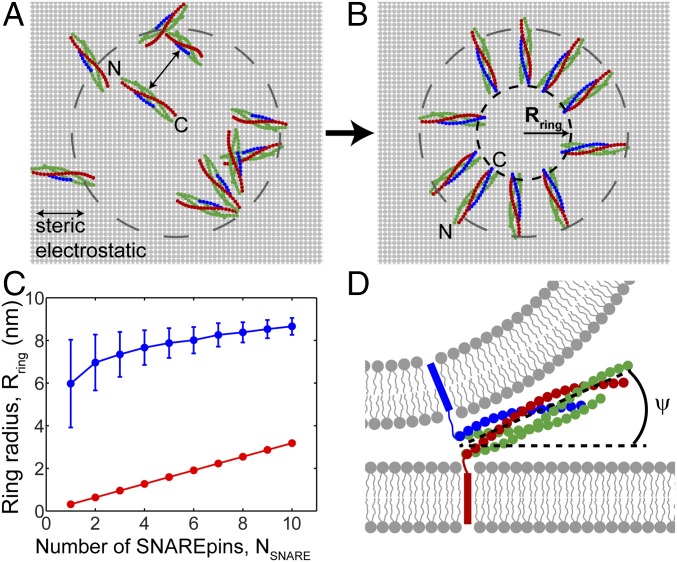
References
-
- Söllner T, et al. SNAP receptors implicated in vesicle targeting and fusion. Nature. 1993;362:318–324. - PubMed
-
- Weber T, et al. SNAREpins: Minimal machinery for membrane fusion. Cell. 1998;92:759–772. - PubMed
-
- Jahn R, Scheller RH. SNAREs–Engines for membrane fusion. Nat Rev Mol Cell Biol. 2006;7:631–643. - PubMed
-
- Lentz BR, Lee JK. Poly(ethylene glycol) (PEG)-mediated fusion between pure lipid bilayers: A mechanism in common with viral fusion and secretory vesicle release? Mol Membr Biol. 1999;16:279–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
