

Nonlinear trade-offs allow the cooperation game to evolve from Prisoner's Dilemma to Snowdrift
- PMID: 28490625
- PMCID: PMC5443946
- DOI: 10.1098/rspb.2017.0228
Nonlinear trade-offs allow the cooperation game to evolve from Prisoner's Dilemma to Snowdrift
Abstract
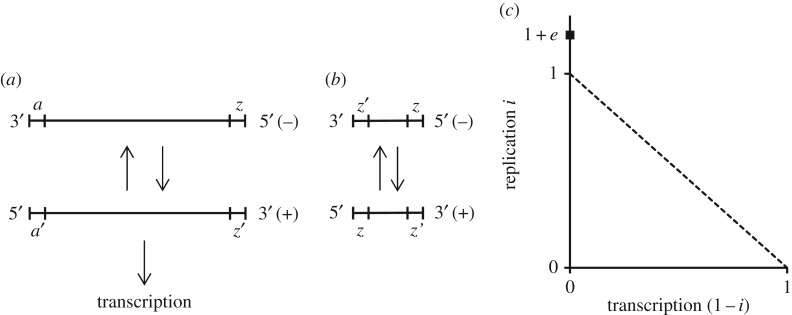
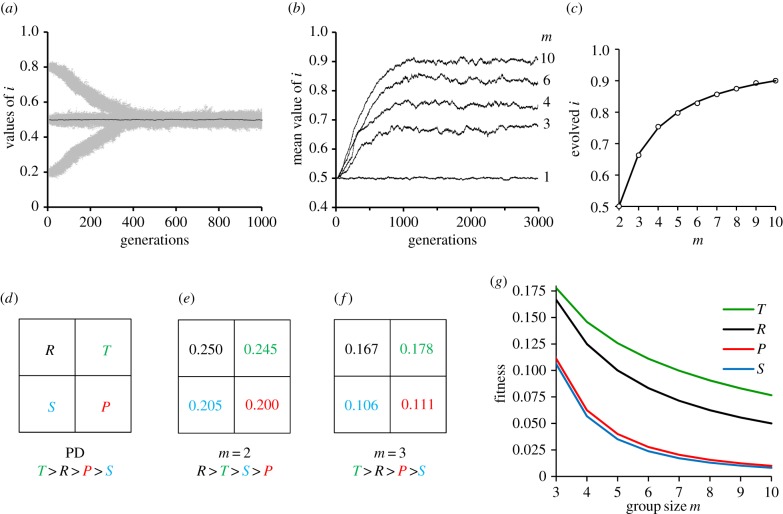
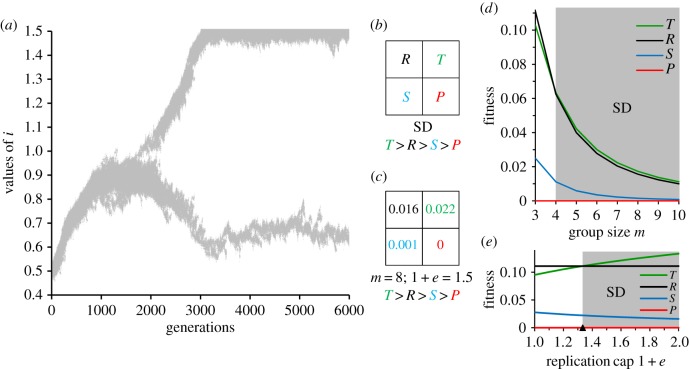
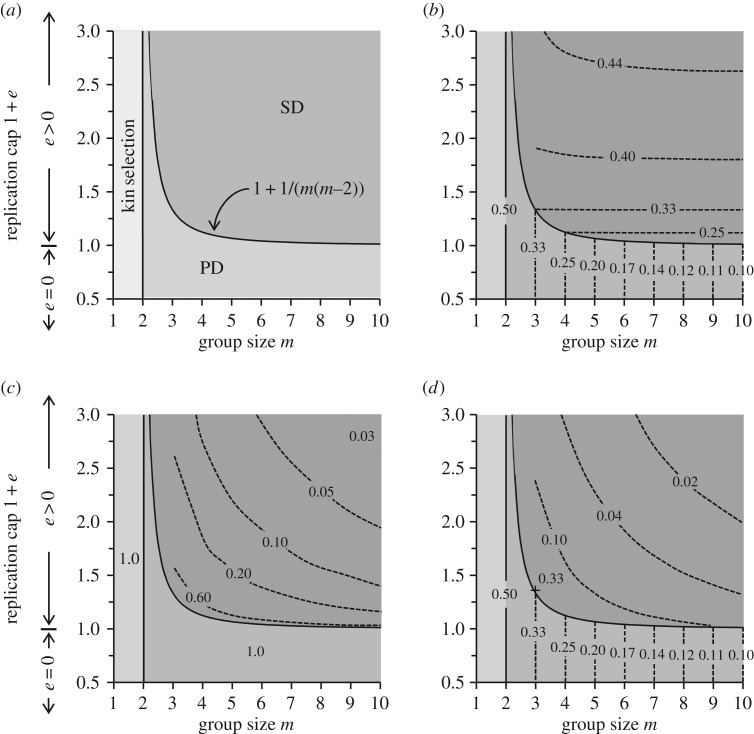
The existence of cooperation, or the production of public goods, is an evolutionary problem. Cooperation is not favoured because the Prisoner's Dilemma (PD) game drives cooperators to extinction. We have re-analysed this problem by using RNA viruses to motivate a model for the evolution of cooperation. Gene products are the public goods and group size is the number of virions co-infecting the same host cell. Our results show that if the trade-off between replication and production of gene products is linear, PD is observed. However, if the trade-off is nonlinear, the viruses evolve into separate lineages of ultra-defectors and ultra-cooperators as group size is increased. The nonlinearity was justified by the existence of real viral ultra-defectors, known as defective interfering particles, which gain a nonlinear advantage by being smaller. The evolution of ultra-defectors and ultra-cooperators creates the Snowdrift game, which promotes high-level production of public goods.
Keywords: Prisoner's Dilemma; RNA viruses; Snowdrift; cooperation; defective interfering particles; game theory.
© 2017 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
References
-
- Doebeli M, Hauert C. 2005. Models of cooperation based on the Prisoner's Dilemma and the Snowdrift game. Ecol. Lett. 8, 748–766. ( 10.1111/j.1461-0248.2005.00773.x) - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources