

TXNIP regulates mitophagy in retinal Müller cells under high-glucose conditions: implications for diabetic retinopathy
- PMID: 28492550
- PMCID: PMC5520711
- DOI: 10.1038/cddis.2017.190
TXNIP regulates mitophagy in retinal Müller cells under high-glucose conditions: implications for diabetic retinopathy
Abstract
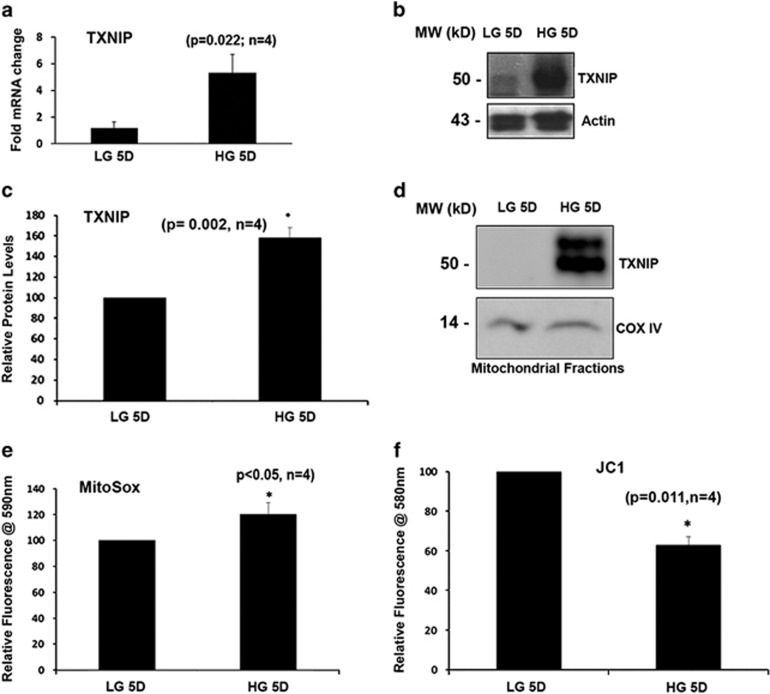
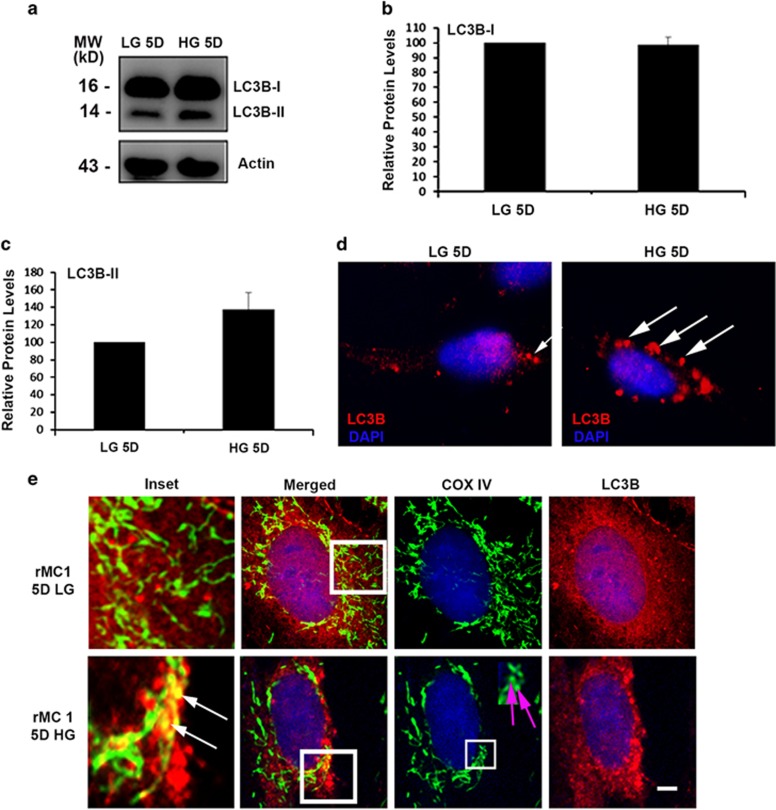
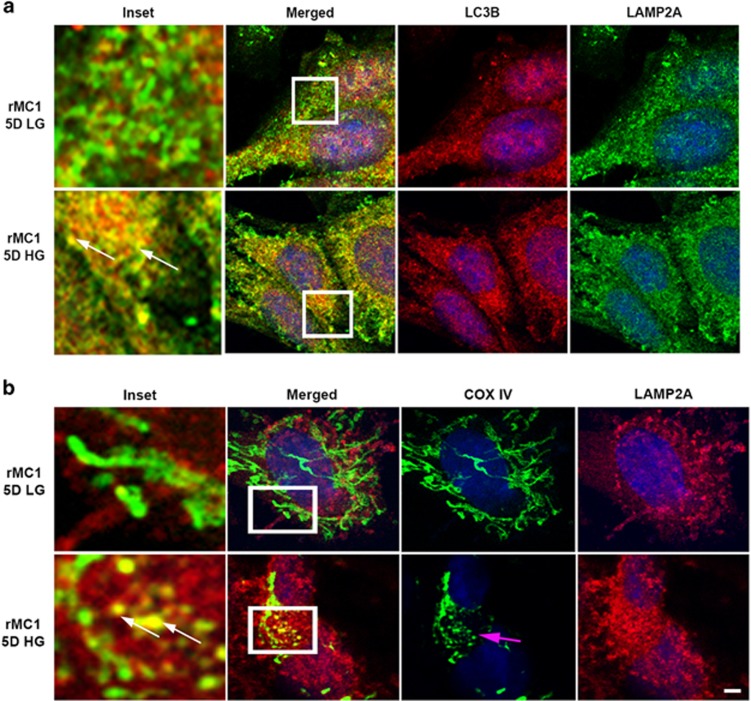
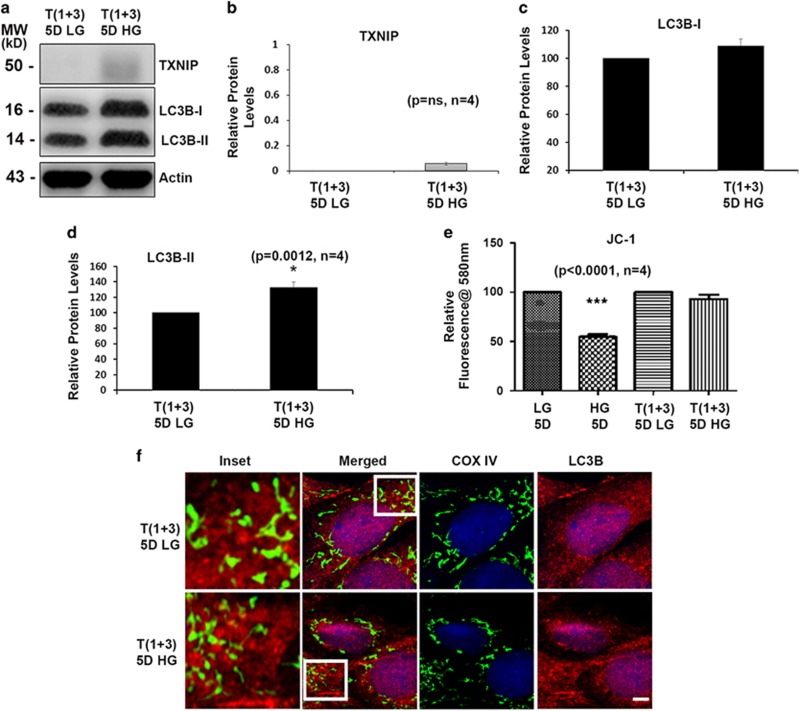
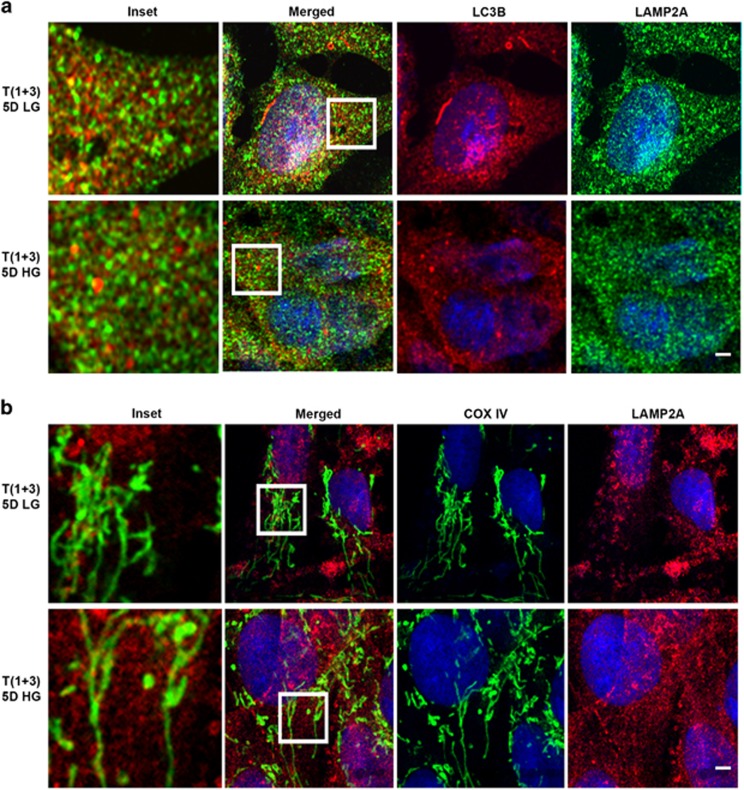
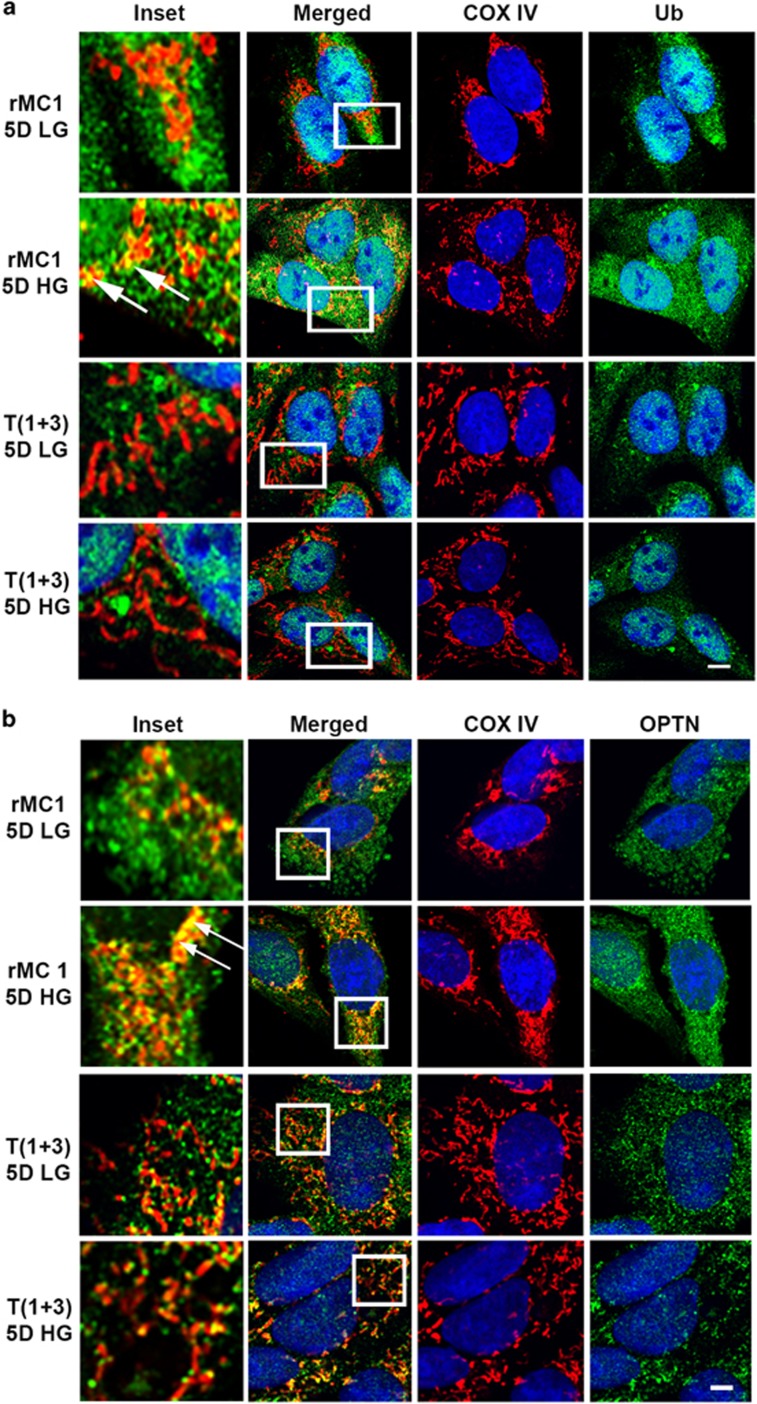
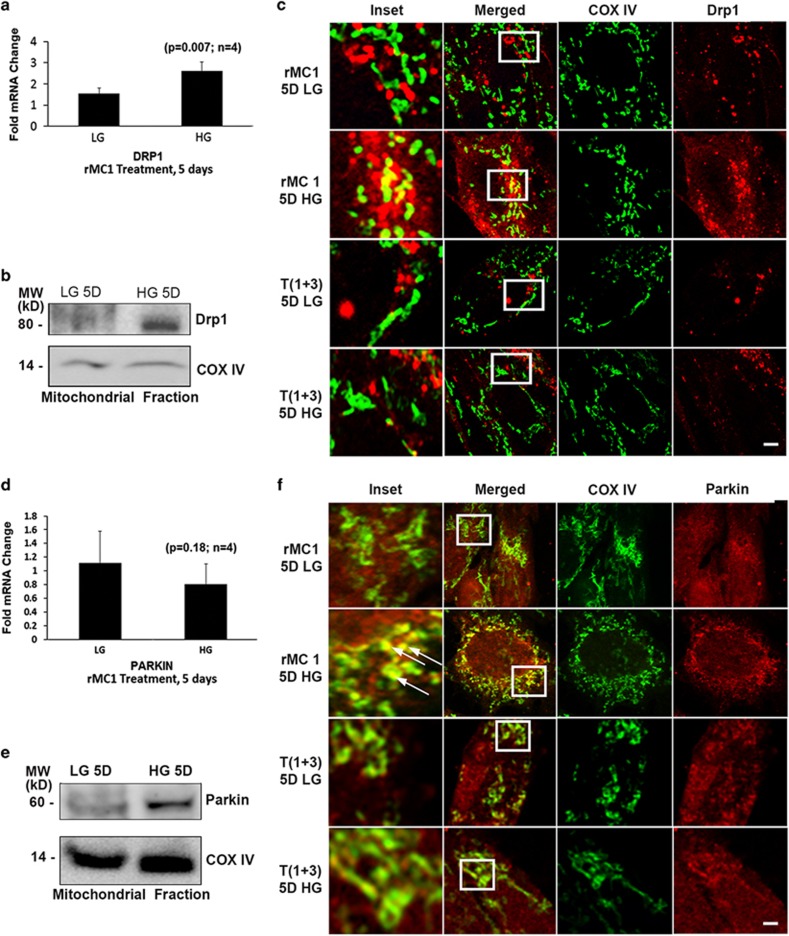
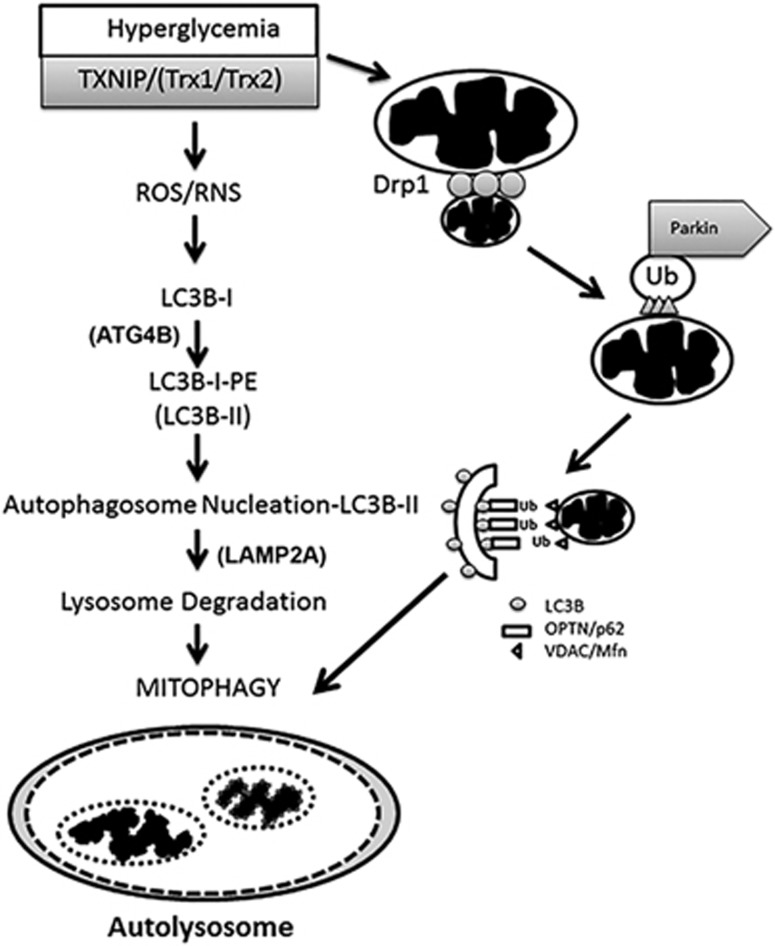
Thioredoxin-interacting protein (TXNIP) is involved in oxidative stress and apoptosis in diabetic retinopathy. However, the role of TXNIP in the removal of damaged mitochondria (MT) via mitophagy, a process of macroautophagy, remains unexplored. Here we investigate the associated cellular and molecular mechanisms underlying mitophagy in retinal cells under diabetic conditions. For this, we maintained a rat Müller cell line (rMC1) under high-glucose (25 mM, HG) or low-glucose (5.5 mM, LG) condition for 5 days. Our data reveal that HG upregulates TXNIP in the cytosol as well as in the MT. Moreover, mitochondrial oxidative stress and membrane depolarization occur under prolonged hyperglycemia leading to fragmentation. These damaged MT are targeted to lysosome for mitophagic degradation, as is evident by co-localization of mitochondrial protein COXIV, a subunit of cytochrome c oxidase, with autophagosome marker LC3BII and the lysosomal membrane protein LAMP2A. In addition, under HG conditions, there is an accumulation of dynamin-related fission protein Drp1 and E3 ubiquitin ligase Parkin in damaged MT, suggesting their roles in mitochondrial fragmentation and ubiquitination, respectively, which is absent in LG conditions. Subsequently, ubiquitin receptors, optineurin and p62/sequestrome 1, bind to the damaged MT and target them to LC3BII autophagosomes. Conversely, TXNIP knockout via CRISPR/Cas9 and TXNIP gRNA prevents the HG-induced mitochondrial damage and mitophagy in rMC1. Last, TXNIP level is also significantly upregulated in the diabetic rat retina in vivo and induces radial glial fibrillary acidic protein expression, a marker for Müller glia activation, and the formation of LC3BII puncta, which are prevented by intravitreal injection of TXNIP siRNA. Therefore, TXNIP represents a potential target for preventing ocular complications of diabetes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Potential Combination Drug Therapy to Prevent Redox Stress and Mitophagy Dysregulation in Retinal Müller Cells under High Glucose Conditions: Implications for Diabetic Retinopathy.Diseases. 2021 Dec 14;9(4):91. doi: 10.3390/diseases9040091. Diseases. 2021. PMID: 34940029 Free PMC article.
-
TXNIP positively regulates the autophagy and apoptosis in the rat müller cell of diabetic retinopathy.Life Sci. 2021 Feb 15;267:118988. doi: 10.1016/j.lfs.2020.118988. Epub 2021 Jan 4. Life Sci. 2021. PMID: 33412212
-
TXNIP links innate host defense mechanisms to oxidative stress and inflammation in retinal Muller glia under chronic hyperglycemia: implications for diabetic retinopathy.Exp Diabetes Res. 2012;2012:438238. doi: 10.1155/2012/438238. Epub 2012 Mar 18. Exp Diabetes Res. 2012. PMID: 22474421 Free PMC article.
-
Mitochondrial dysfunction and endoplasmic reticulum stress in diabetic retinopathy: mechanistic insights into high glucose-induced retinal cell death.Curr Clin Pharmacol. 2013 Nov;8(4):278-84. doi: 10.2174/1574884711308040003. Curr Clin Pharmacol. 2013. PMID: 23173958 Review.
-
The role of Müller cell glucocorticoid signaling in diabetic retinopathy.Graefes Arch Clin Exp Ophthalmol. 2020 Feb;258(2):221-230. doi: 10.1007/s00417-019-04521-w. Epub 2019 Nov 16. Graefes Arch Clin Exp Ophthalmol. 2020. PMID: 31734719 Review.
Cited by
-
MiR-17-5p Inhibits TXNIP/NLRP3 Inflammasome Pathway and Suppresses Pancreatic β-Cell Pyroptosis in Diabetic Mice.Front Cardiovasc Med. 2021 Nov 22;8:768029. doi: 10.3389/fcvm.2021.768029. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 34881312 Free PMC article.
-
The Role of Txnip in Mitophagy Dysregulation and Inflammasome Activation in Diabetic Retinopathy: A New Perspective.JOJ Ophthalmol. 2017;4(4):10.19080/jojo.2017.04.555643. doi: 10.19080/jojo.2017.04.555643. Epub 2017 Sep 15. JOJ Ophthalmol. 2017. PMID: 29376145 Free PMC article.
-
Thioredoxin-interacting protein in diabetic retinal neurodegeneration: A novel potential therapeutic target for diabetic retinopathy.Front Neurosci. 2022 Aug 9;16:957667. doi: 10.3389/fnins.2022.957667. eCollection 2022. Front Neurosci. 2022. PMID: 36017183 Free PMC article. Review.
-
Early impairments in the retina of rats fed with high fructose/high fat diet are associated with glucose metabolism deregulation but not dyslipidaemia.Sci Rep. 2019 Apr 12;9(1):5997. doi: 10.1038/s41598-019-42528-9. Sci Rep. 2019. PMID: 30979946 Free PMC article.
-
TXNIP, a novel key factor to cause Schwann cell dysfunction in diabetic peripheral neuropathy, under the regulation of PI3K/Akt pathway inhibition-induced DNMT1 and DNMT3a overexpression.Cell Death Dis. 2021 Jun 23;12(7):642. doi: 10.1038/s41419-021-03930-2. Cell Death Dis. 2021. PMID: 34162834 Free PMC article.
References
-
- Perrone L, Devi TS, Hosoya K, Terasaki T, Singh LP. Thioredoxin interacting protein (TXNIP) induces inflammation through chromatin modification in retinal capillary endothelial cells under diabetic conditions. J Cell Physiol 2009; 221: 262–272. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
