

Selective pre-priming of HA-specific CD4 T cells restores immunological reactivity to HA on heterosubtypic influenza infection
- PMID: 28493882
- PMCID: PMC5426616
- DOI: 10.1371/journal.pone.0176407
Selective pre-priming of HA-specific CD4 T cells restores immunological reactivity to HA on heterosubtypic influenza infection
Abstract
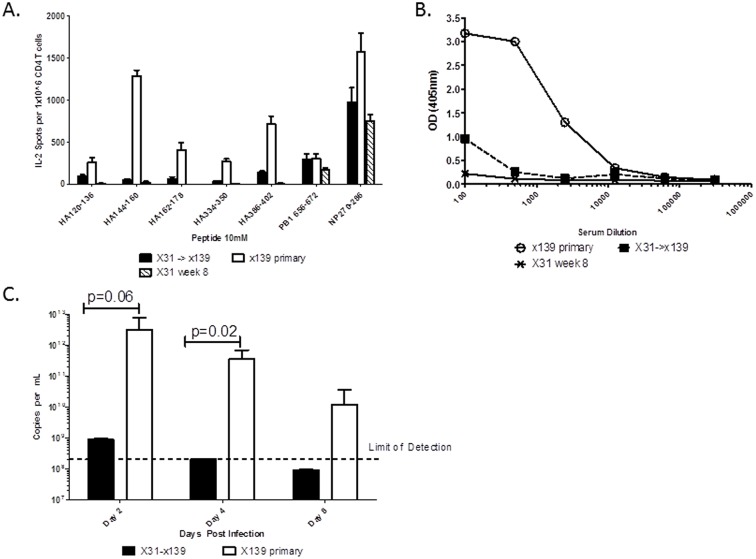
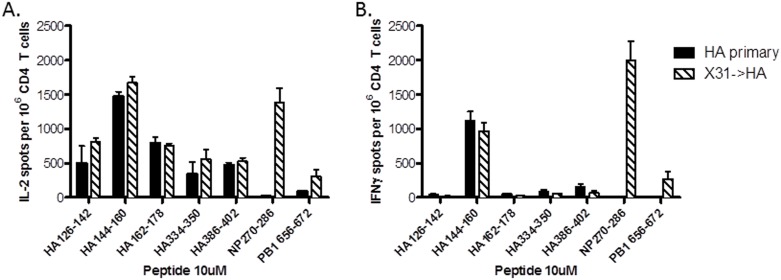
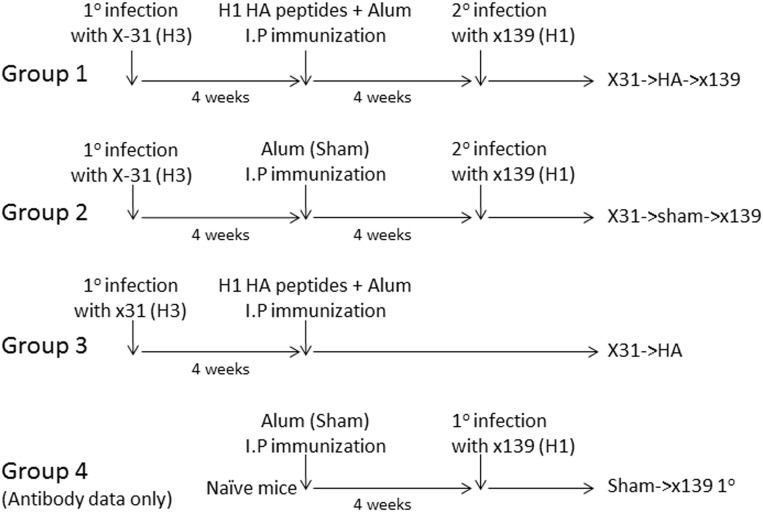
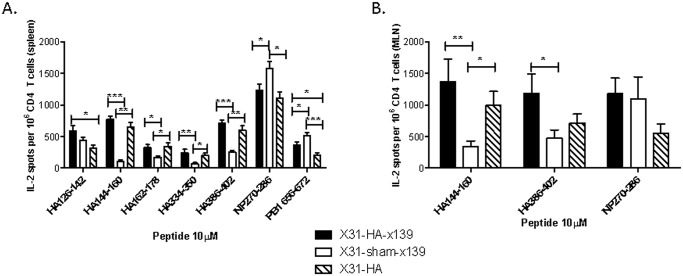
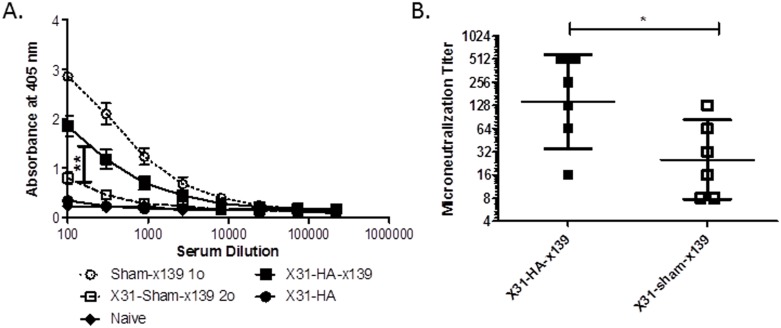
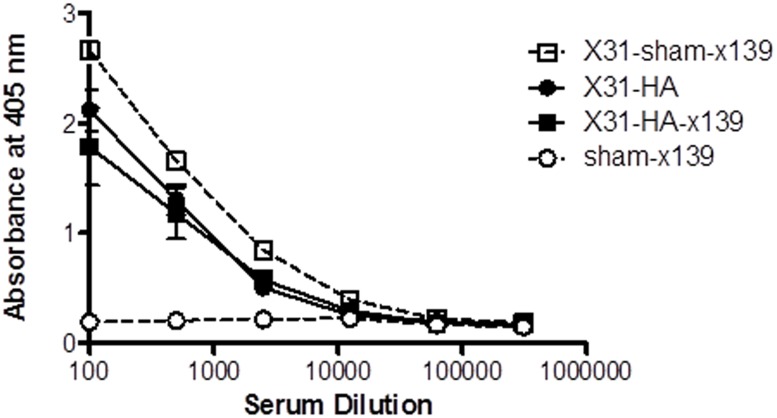
A hallmark of the immune response to influenza is repeated encounters with proteins containing both genetically conserved and variable components. Therefore, the B and T cell repertoire is continually being remodeled, with competition between memory and naïve lymphocytes. Our previous work using a mouse model of secondary heterosubtypic influenza infection has shown that this competition results in a focusing of CD4 T cell response specificity towards internal virion proteins with a selective decrease in CD4 T cell reactivity to the novel HA epitopes. Strikingly, this shift in CD4 T cell specificity was associated with a diminished anti-HA antibody response. Here, we sought to determine whether the loss in HA-specific reactivity that occurs as a consequence of immunological memory could be reversed by selectively priming HA-specific CD4 T cells prior to secondary infection. Using a peptide-based priming strategy, we found that selective expansion of the anti-HA CD4 T cell memory repertoire enhanced HA-specific antibody production upon heterosubtypic infection. These results suggest that the potentially deleterious consequences of repeated exposure to conserved influenza internal virion proteins could be reversed by vaccination strategies that selectively arm the HA-specific CD4 T cell compartment. This could be a potentially useful pre-pandemic vaccination strategy to promote accelerated neutralizing antibody production on challenge with a pandemic influenza strain that contains few conserved HA epitopes.
Conflict of interest statement
Figures
Similar articles
-
Hemagglutinin-specific CD4+ T-cell responses following 2009-pH1N1 inactivated split-vaccine inoculation in humans.Vaccine. 2017 Oct 9;35(42):5644-5652. doi: 10.1016/j.vaccine.2017.08.061. Epub 2017 Sep 13. Vaccine. 2017. PMID: 28917539 Clinical Trial.
-
CD4+ T-cell expansion predicts neutralizing antibody responses to monovalent, inactivated 2009 pandemic influenza A(H1N1) virus subtype H1N1 vaccine.J Infect Dis. 2013 Jan 15;207(2):297-305. doi: 10.1093/infdis/jis684. Epub 2012 Nov 12. J Infect Dis. 2013. PMID: 23148285 Free PMC article.
-
Cutting edge: Heterosubtypic influenza infection antagonizes elicitation of immunological reactivity to hemagglutinin.J Immunol. 2013 Aug 1;191(3):1001-5. doi: 10.4049/jimmunol.1203520. Epub 2013 Jun 21. J Immunol. 2013. PMID: 23794632 Free PMC article.
-
Recalling the Future: Immunological Memory Toward Unpredictable Influenza Viruses.Front Immunol. 2019 Jul 2;10:1400. doi: 10.3389/fimmu.2019.01400. eCollection 2019. Front Immunol. 2019. PMID: 31312199 Free PMC article. Review.
-
Keeping the memory of influenza viruses.Pathol Biol (Paris). 2010 Apr;58(2):e79-86. doi: 10.1016/j.patbio.2010.01.010. Epub 2010 Mar 19. Pathol Biol (Paris). 2010. PMID: 20303671 Review.
Cited by
-
Harnessing the Power of T Cells: The Promising Hope for a Universal Influenza Vaccine.Vaccines (Basel). 2018 Mar 26;6(2):18. doi: 10.3390/vaccines6020018. Vaccines (Basel). 2018. PMID: 29587436 Free PMC article. Review.
-
CD4 T cell epitope abundance in ferritin core potentiates responses to hemagglutinin nanoparticle vaccines.NPJ Vaccines. 2022 Oct 26;7(1):124. doi: 10.1038/s41541-022-00547-0. NPJ Vaccines. 2022. PMID: 36289232 Free PMC article.
-
Impact of antigenic evolution and original antigenic sin on SARS-CoV-2 immunity.J Clin Invest. 2023 Jan 3;133(1):e162192. doi: 10.1172/JCI162192. J Clin Invest. 2023. PMID: 36594464 Free PMC article. Review.
-
CD4 T cells in protection from influenza virus: Viral antigen specificity and functional potential.Immunol Rev. 2018 Jul;284(1):91-105. doi: 10.1111/imr.12662. Immunol Rev. 2018. PMID: 29944766 Free PMC article. Review.
-
Distinct and complementary roles of CD4 T cells in protective immunity to influenza virus.Curr Opin Immunol. 2018 Aug;53:13-21. doi: 10.1016/j.coi.2018.03.019. Epub 2018 Apr 2. Curr Opin Immunol. 2018. PMID: 29621639 Free PMC article. Review.
References
-
- Thompson WW, Shay DK, Weintraub E, Brammer L, Cox N, Anderson LJ, et al. (2003) Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA 289: 179–186. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
