

Ezrin interacts with S100A4 via both its N- and C-terminal domains
- PMID: 28493957
- PMCID: PMC5426754
- DOI: 10.1371/journal.pone.0177489
Ezrin interacts with S100A4 via both its N- and C-terminal domains
Abstract
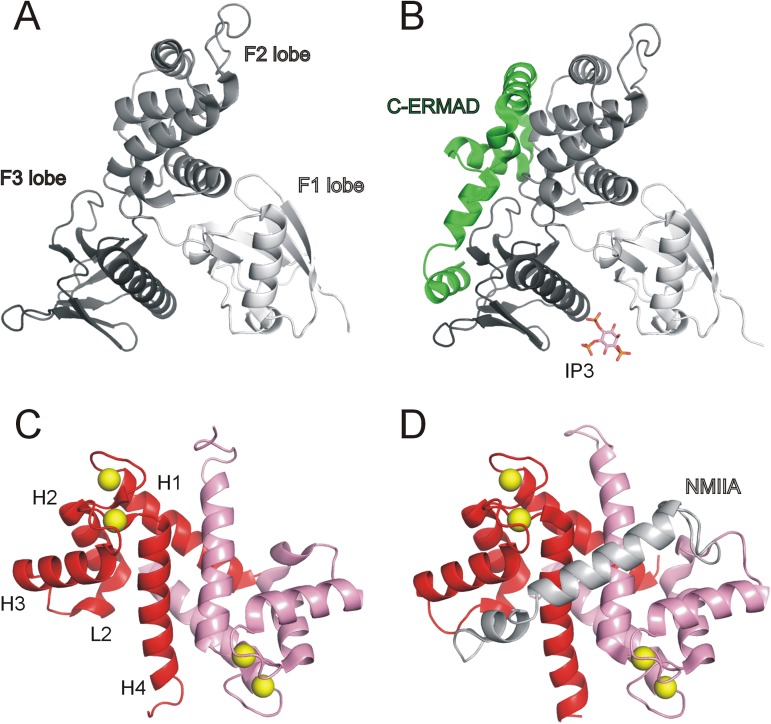
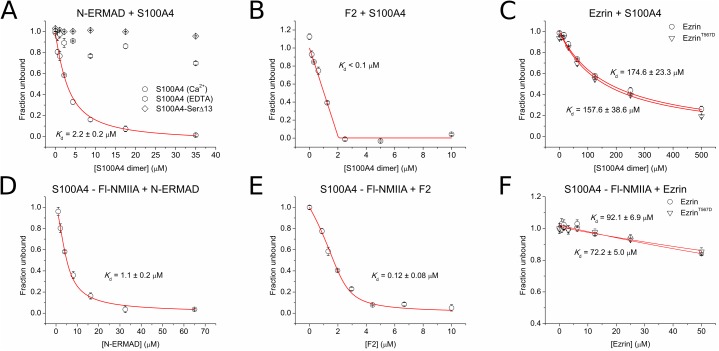
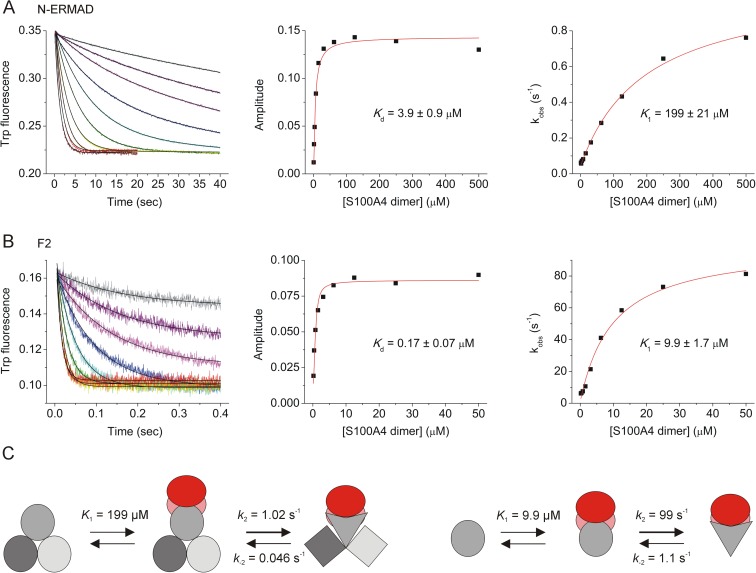
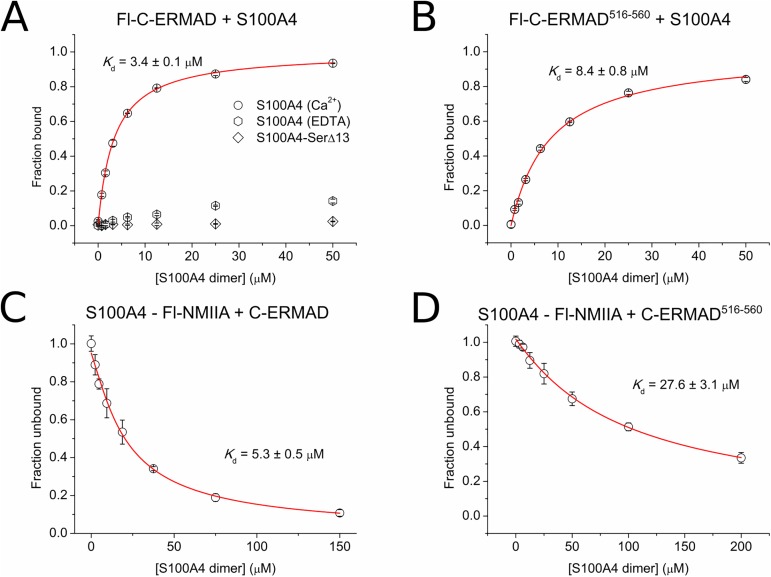
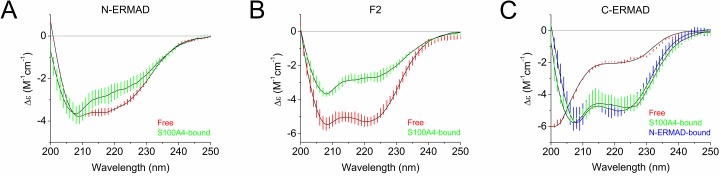
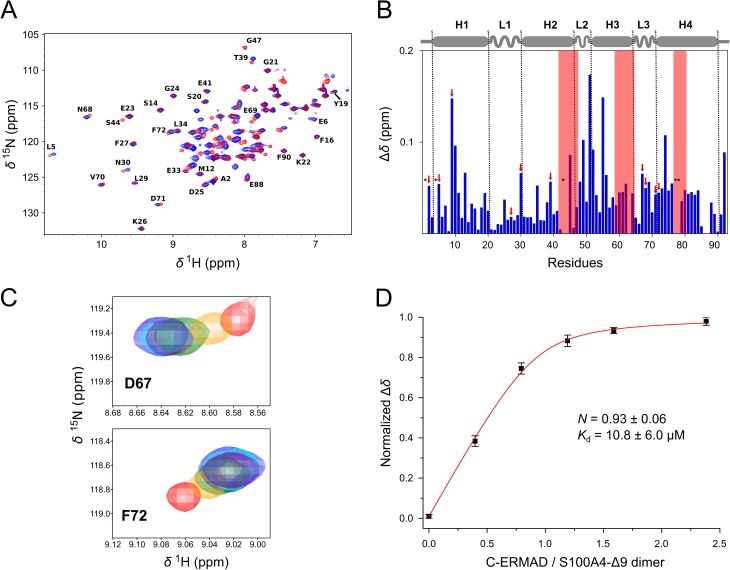
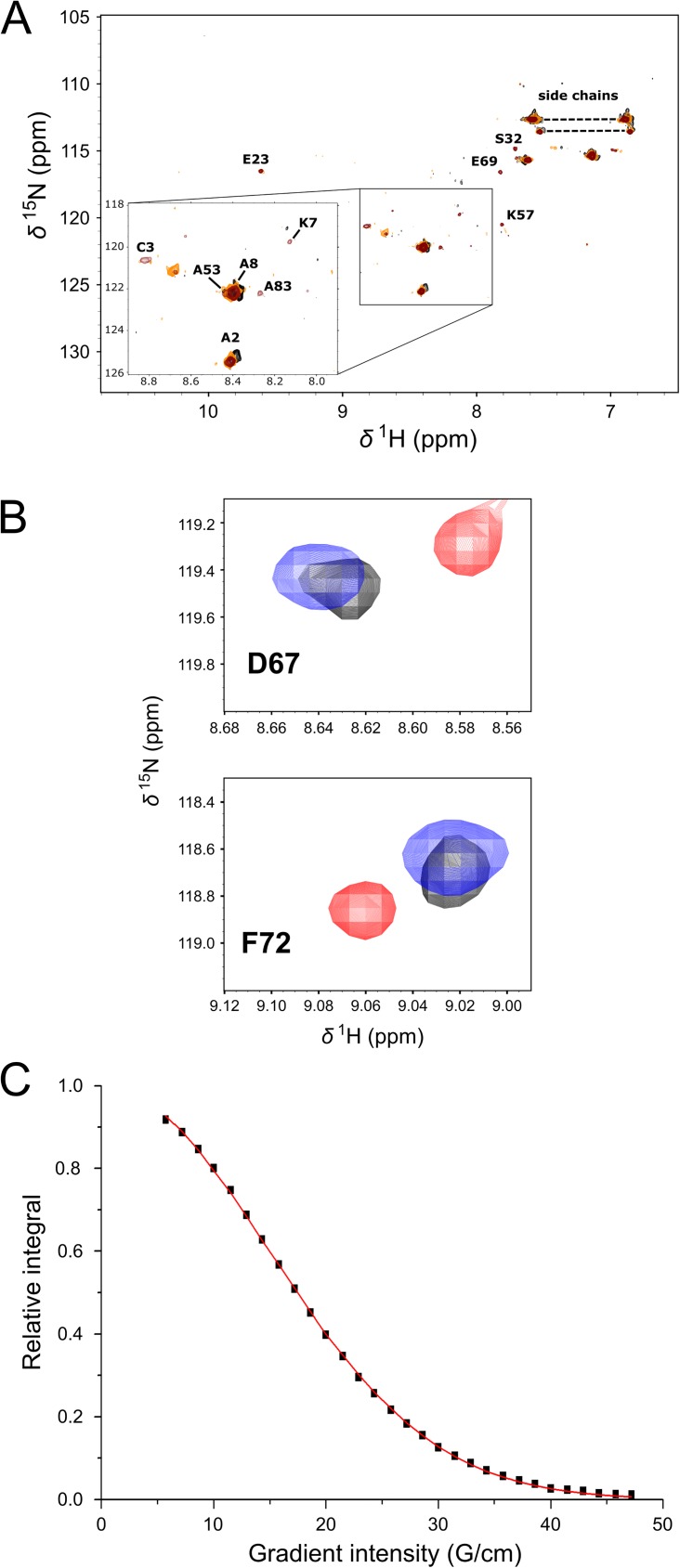
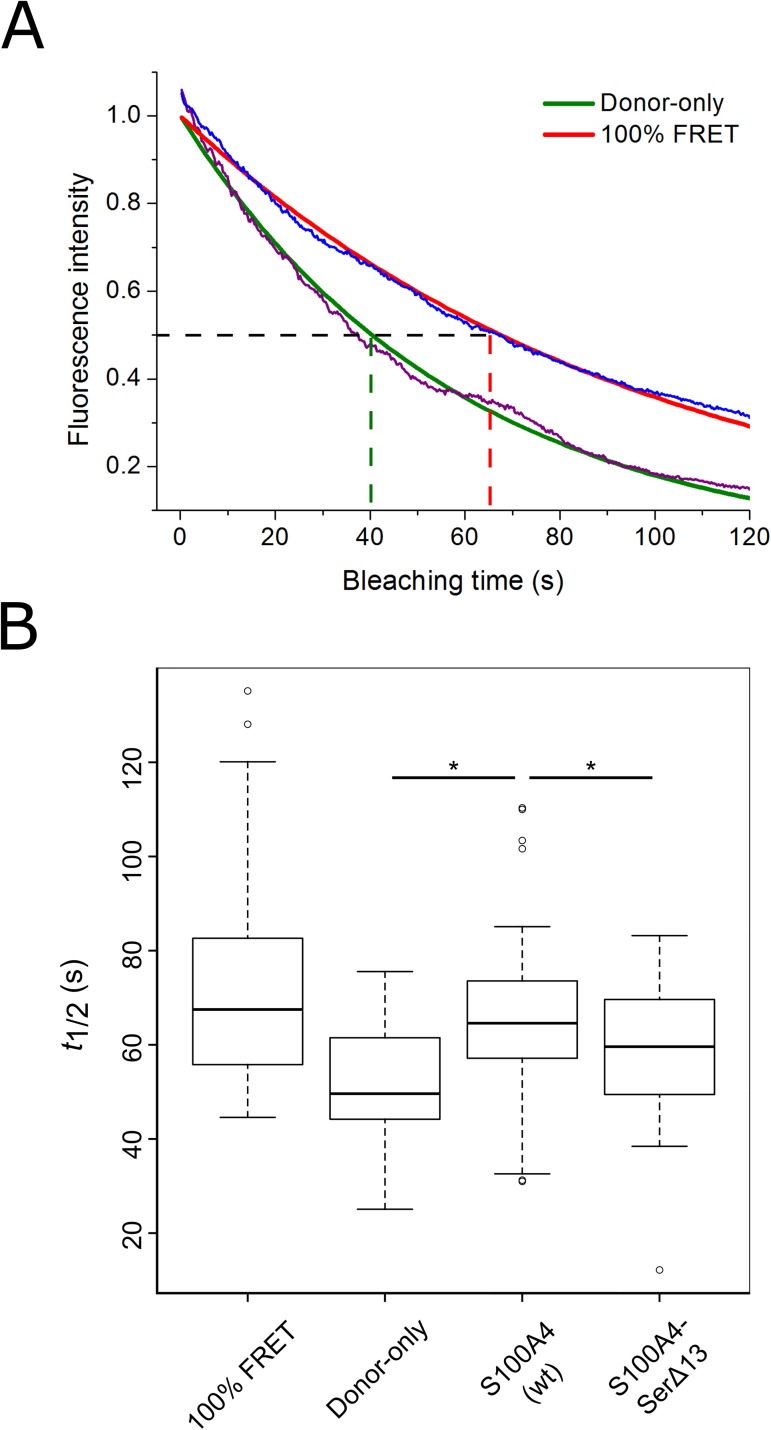
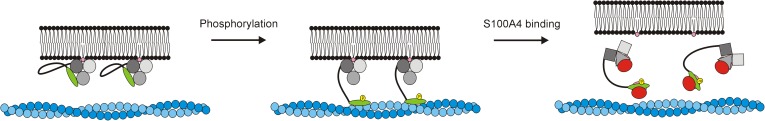
Ezrin belongs to the ERM (ezrin, radixin, moesin) protein family that has a role in cell morphology changes, adhesion and migration as an organizer of the cortical cytoskeleton by linking actin filaments to the apical membrane of epithelial cells. It is highly expressed in a variety of human cancers and promotes metastasis. Members of the Ca2+-binding EF-hand containing S100 proteins have similar pathological properties; they are overexpressed in cancer cells and involved in metastatic processes. In this study, using tryptophan fluorescence and stopped-flow kinetics, we show that S100A4 binds to the N-terminal ERM domain (N-ERMAD) of ezrin with a micromolar affinity. The binding involves the F2 lobe of the N-ERMAD and follows an induced fit kinetic mechanism. Interestingly, S100A4 binds also to the unstructured C-terminal actin binding domain (C-ERMAD) with similar affinity. Using NMR spectroscopy, we characterized the complex of S100A4 with the C-ERMAD and demonstrate that no ternary complex is simultaneously formed with the two ezrin domains. Furthermore, we show that S100A4 co-localizes with ezrin in HEK-293T cells. However, S100A4 very weakly binds to full-length ezrin in vitro indicating that the interaction of S100A4 with ezrin requires other regulatory events such as protein phosphorylation and/or membrane binding, shifting the conformational equilibrium of ezrin towards the open state. As both proteins play an important role in promoting metastasis, the characterization of their interaction could shed more light on the molecular events contributing to this pathological process.
Conflict of interest statement
Figures
References
-
- Neisch AL, Fehon RG. Ezrin, Radixin and Moesin: key regulators of membrane-cortex interactions and signaling. Curr Opin Cell Biol. 2011;23(4):377–82. Epub 2011/05/20. PubMed Central PMCID: PMC3148288. doi: 10.1016/j.ceb.2011.04.011 - DOI - PMC - PubMed
-
- Clucas J, Valderrama F. ERM proteins in cancer progression. J Cell Sci. 2015;128(6):1253 Epub 2015/03/17. doi: 10.1242/jcs.170027 - DOI - PubMed
-
- Barreiro O, Yanez-Mo M, Serrador JM, Montoya MC, Vicente-Manzanares M, Tejedor R, et al. Dynamic interaction of VCAM-1 and ICAM-1 with moesin and ezrin in a novel endothelial docking structure for adherent leukocytes. The Journal of cell biology. 2002;157(7):1233–45. PubMed Central PMCID: PMC2173557. doi: 10.1083/jcb.200112126 - DOI - PMC - PubMed
-
- Granes F, Urena JM, Rocamora N, Vilaro S. Ezrin links syndecan-2 to the cytoskeleton. J Cell Sci. 2000;113 (Pt 7):1267–76. - PubMed
-
- Martin TA, Harrison G, Mansel RE, Jiang WG. The role of the CD44/ezrin complex in cancer metastasis. Critical reviews in oncology/hematology. 2003;46(2):165–86. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
