

Environmental Calcium Controls Alternate Physical States of the Caulobacter Surface Layer
- PMID: 28494955
- PMCID: PMC5425405
- DOI: 10.1016/j.bpj.2017.04.003
Environmental Calcium Controls Alternate Physical States of the Caulobacter Surface Layer
Abstract
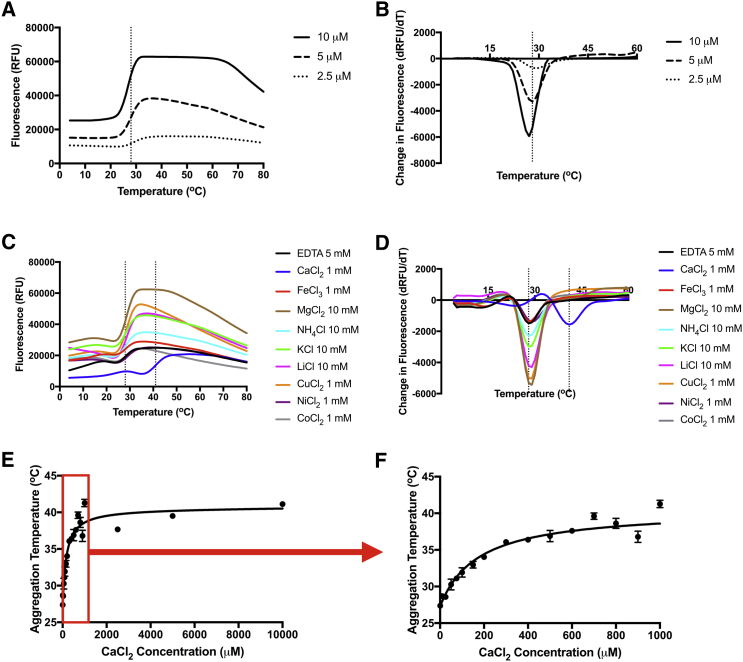
Surface layers (S-layers) are paracrystalline, proteinaceous structures found in most archaea and many bacteria. Often the outermost cell envelope component, S-layers serve diverse functions including aiding pathogenicity and protecting against predators. We report that the S-layer of Caulobacter crescentus exhibits calcium-mediated structural plasticity, switching irreversibly between an amorphous aggregate state and the crystalline state. This finding invalidates the common assumption that S-layers serve only as static wall-like structures. In vitro, the Caulobacter S-layer protein, RsaA, enters the aggregate state at physiological temperatures and low divalent calcium ion concentrations. At higher concentrations, calcium ions stabilize monomeric RsaA, which can then transition to the two-dimensional crystalline state. Caulobacter requires micromolar concentrations of calcium for normal growth and development. Without an S-layer, Caulobacter is even more sensitive to changes in environmental calcium concentration. Therefore, this structurally dynamic S-layer responds to environmental conditions as an ion sensor and protects Caulobacter from calcium deficiency stress, a unique mechanism of bacterial adaptation. These findings provide a biochemical and physiological basis for RsaA's calcium-binding behavior, which extends far beyond calcium's commonly accepted role in aiding S-layer biogenesis or oligomerization and demonstrates a connection to cellular fitness.
Copyright © 2017 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures







References
-
- Albers S.-V., Meyer B.H. The archaeal cell envelope. Nat. Rev. Microbiol. 2011;9:414–426. - PubMed
-
- Sleytr U.B., Sára M. Bacterial and archaeal S-layer proteins: structure-function relationships and their biotechnological applications. Trends Biotechnol. 1997;15:20–26. - PubMed
-
- Gerbino E., Carasi P., Gómez-Zavaglia A. Role of S-layer proteins in bacteria. World J. Microbiol. Biotechnol. 2015;31:1877–1887. - PubMed
-
- Sleytr U.B., Messner P., Sára M. Crystalline bacterial cell surface layers. Mol. Microbiol. 1993;10:911–916. - PubMed
-
- Fagan R.P., Fairweather N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 2014;12:211–222. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
