

Translating in vitro ligand bias into in vivo efficacy
- PMID: 28495495
- PMCID: PMC5675828
- DOI: 10.1016/j.cellsig.2017.05.002
Translating in vitro ligand bias into in vivo efficacy
Abstract
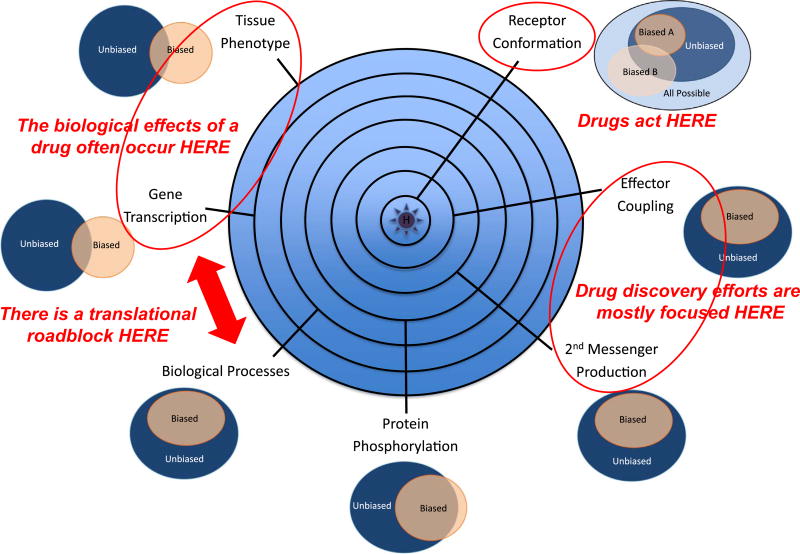
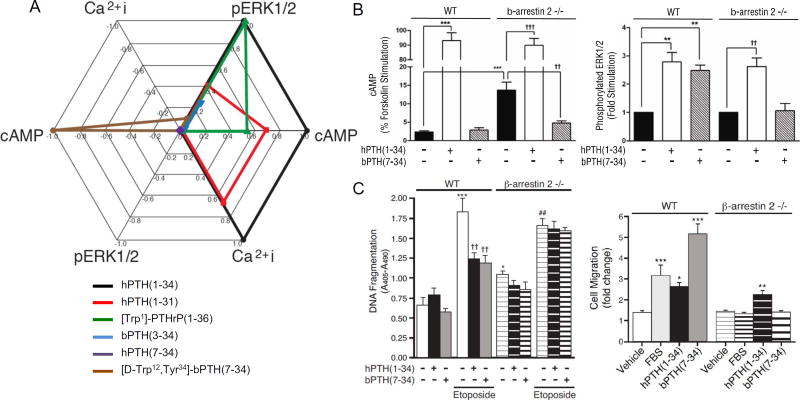
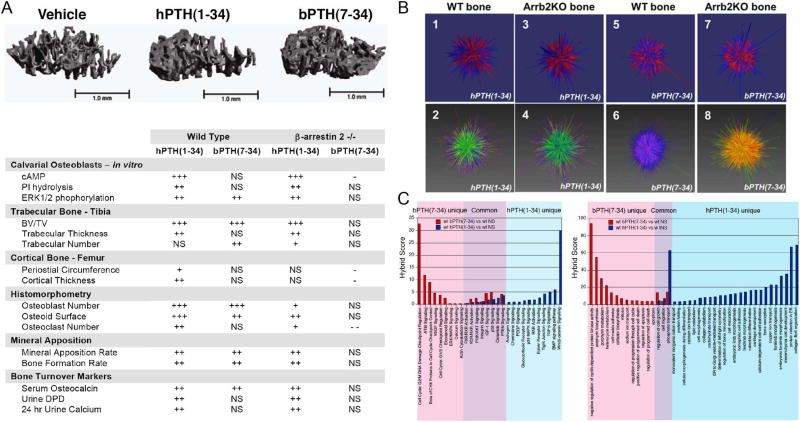
It is increasingly apparent that ligand structure influences both the efficiency with which G protein-coupled receptors (GPCRs) engage their downstream effectors and the manner in which they are activated. Thus, 'biased' agonists, synthetic ligands whose intrinsic efficacy differs from the native ligand, afford a strategy for manipulating GPCR signaling in ways that promote beneficial signals while blocking potentially deleterious ones. Still, there are significant challenges in relating in vitro ligand efficacy, which is typically measured in heterologous expression systems, to the biological response in vivo, where the ligand is acting on natively expressed receptors and in the presence of the endogenous ligand. This is particularly true of arrestin pathway-selective 'biased' agonists. The type 1 parathyroid hormone receptor (PTH1R) is a case in point. Parathyroid hormone (PTH) is the principal physiological regulator of calcium homeostasis, and PTH1R expressed on cells of the osteoblast lineage are an established therapeutic target in osteoporosis. In vitro, PTH1R signaling is highly sensitive to ligand structure, and PTH analogs that affect the selectivity/kinetics of G protein coupling or that engage arrestin-dependent signaling mechanisms without activating heterotrimeric G proteins have been identified. In vivo, intermittent administration of conventional PTH analogs accelerates the rate of osteoblastic bone formation, largely through known cAMP-dependent mechanisms. Paradoxically, both intermittent and continuous administration of an arrestin pathway-selective PTH analog, which in vivo would be expected to antagonize endogenous PTH1R-cAMP signaling, also increases bone mass. Transcriptomic analysis of tissue from treated animals suggests that conventional and arrestin pathway-selective PTH1R ligands act in largely different ways, with the latter principally affecting pathways involved in the regulation of cell cycle, survival, and migration/cytoskeletal dynamics. Such multi-dimensional in vitro and in vivo analyses of ligand bias may provide insights into the physiological roles of non-canonical arrestin-mediated signaling pathways in vivo, and provide a conceptual framework for translating arrestin pathway-selective ligands into viable therapeutics.
Keywords: Arrestin; G protein-coupled receptor; Osteoblast; Osteoporosis; Parathyroid hormone; Pharmacology.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflicts of interest related to the content of this article.
Figures
Similar articles
-
Biased agonism at the parathyroid hormone receptor: a demonstration of functional selectivity in bone metabolism.Mini Rev Med Chem. 2012 Aug;12(9):856-65. doi: 10.2174/138955712800959125. Mini Rev Med Chem. 2012. PMID: 22681253 Review.
-
Biasing the parathyroid hormone receptor: relating in vitro ligand efficacy to in vivo biological activity.Methods Enzymol. 2013;522:229-62. doi: 10.1016/B978-0-12-407865-9.00013-3. Methods Enzymol. 2013. PMID: 23374189
-
Arrestins in bone.Prog Mol Biol Transl Sci. 2013;118:335-58. doi: 10.1016/B978-0-12-394440-5.00013-9. Prog Mol Biol Transl Sci. 2013. PMID: 23764060 Review.
-
β-arrestin-biased agonism at the parathyroid hormone receptor uncouples bone formation from bone resorption.Endocr Metab Immune Disord Drug Targets. 2011 Jun;11(2):112-9. doi: 10.2174/187153011795564151. Endocr Metab Immune Disord Drug Targets. 2011. PMID: 21476967 Review.
-
β-Arrestin-biased signaling of PTH analogs of the type 1 parathyroid hormone receptor.Cell Signal. 2013 Feb;25(2):527-38. doi: 10.1016/j.cellsig.2012.11.012. Epub 2012 Nov 15. Cell Signal. 2013. PMID: 23159578
Cited by
-
Pharmacophore-Model-Based Virtual-Screening Approaches Identified Novel Natural Molecular Candidates for Treating Human Neuroblastoma.Curr Issues Mol Biol. 2022 Oct 13;44(10):4838-4858. doi: 10.3390/cimb44100329. Curr Issues Mol Biol. 2022. PMID: 36286044 Free PMC article.
-
Intersection of the Orphan G Protein-Coupled Receptor, GPR19, with the Aging Process.Int J Mol Sci. 2022 Nov 6;23(21):13598. doi: 10.3390/ijms232113598. Int J Mol Sci. 2022. PMID: 36362387 Free PMC article. Review.
-
Functional consequences of spatial, temporal and ligand bias of G protein-coupled receptors.Nat Rev Nephrol. 2024 Nov;20(11):722-741. doi: 10.1038/s41581-024-00869-3. Epub 2024 Jul 22. Nat Rev Nephrol. 2024. PMID: 39039165 Review.
-
Comparison of the pharmacologic profiles of arginine vasopressin and oxytocin analogs at marmoset, titi monkey, macaque, and human oxytocin receptors.Biomed Pharmacother. 2020 May;125:109832. doi: 10.1016/j.biopha.2020.109832. Epub 2020 Feb 1. Biomed Pharmacother. 2020. PMID: 32018219 Free PMC article.
-
Opioid Analgesics: Rise and Fall of Ligand Biased Signaling and Future Perspectives in the Quest for the Holy Grail.CNS Drugs. 2025 Jun;39(6):565-581. doi: 10.1007/s40263-025-01172-w. Epub 2025 Apr 1. CNS Drugs. 2025. PMID: 40169527 Review.
References
-
- De Lean A, Stadel JM, Lefkowitz RJ. A ternary complex model explains the agonist-specific binding properties of the adenylate cyclase-coupled beta-adrenergic receptor. J. Biol. Chem. 1980;255:7108–7117. - PubMed
-
- Samama P, Cotecchia S, Costa T, Lefkowitz RJ. A mutation-induced activated state of the beta 2-adrenergic receptor. Extending the ternary complex model. J. Biol. Chem. 1993;268:4625–4636. - PubMed
-
- Galandrin S, Oligny-Longpré G, Bouvier M. The evasive nature of drug efficacy: implications for drug discovery. Trends Pharmacol. Sci. 2007;28:423–430. - PubMed
-
- Christopoulos A, Kenakin T. G protein-coupled receptor allosterism and complexing. Pharmacol. Rev. 2002;54:323–374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
