

Modeling glucose metabolism and lactate production in the kidney
- PMID: 28495544
- PMCID: PMC5533195
- DOI: 10.1016/j.mbs.2017.04.008
Modeling glucose metabolism and lactate production in the kidney
Abstract
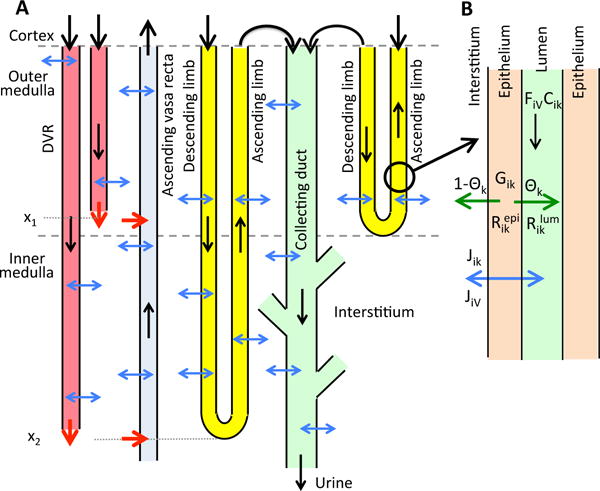
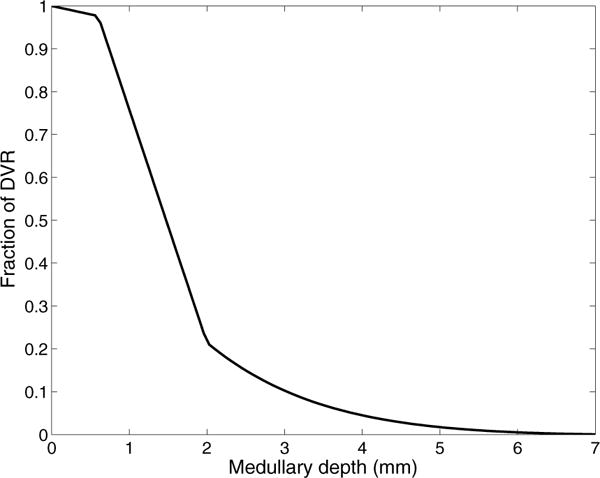
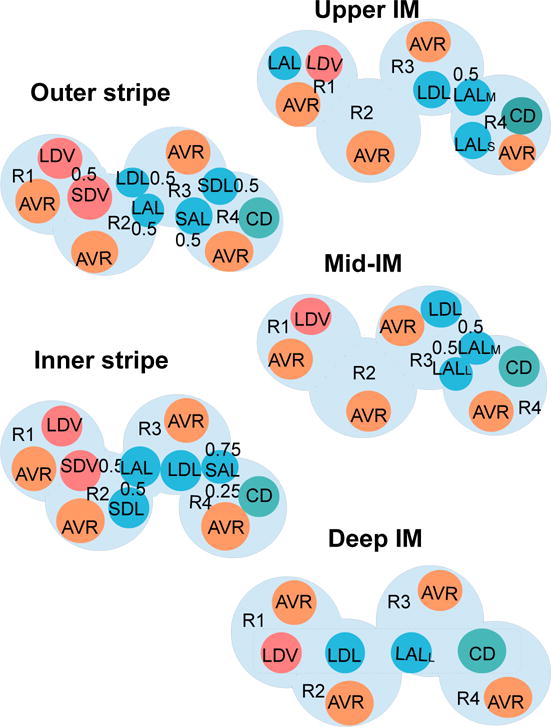
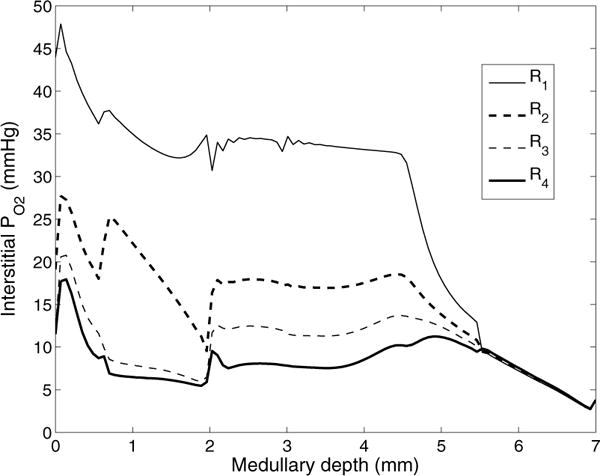
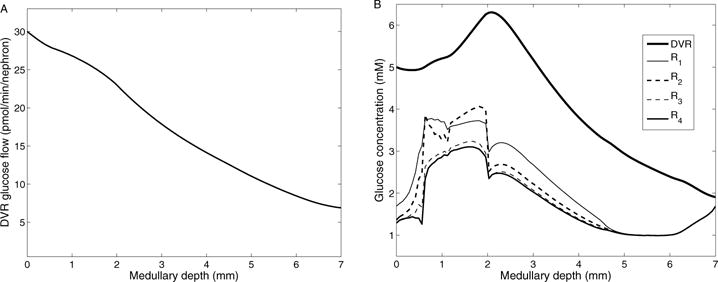
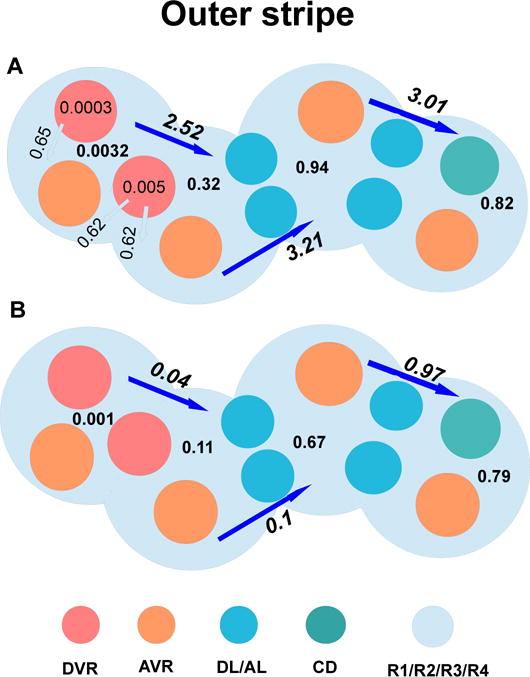
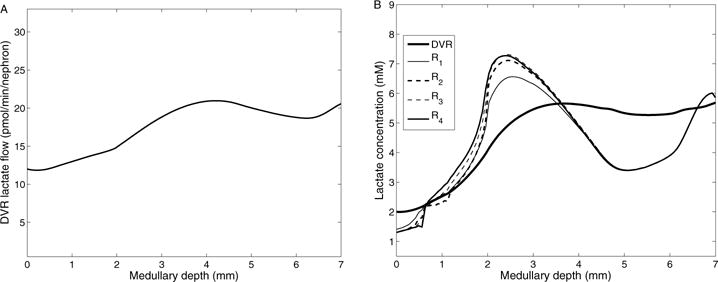
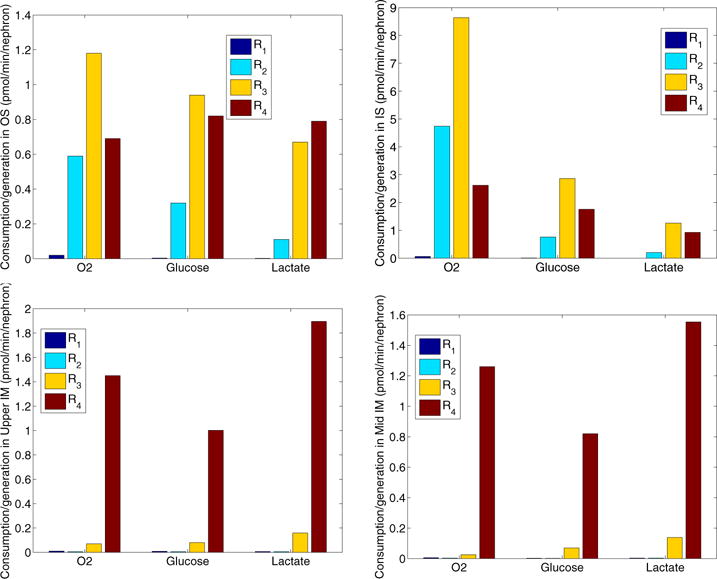
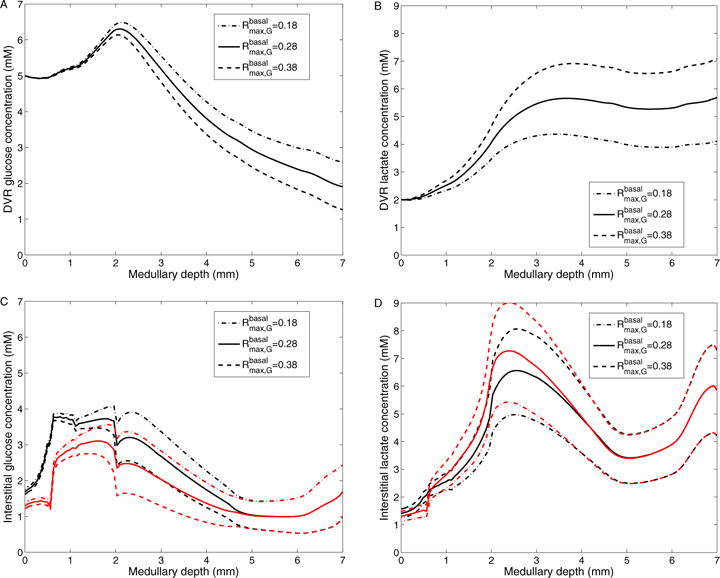
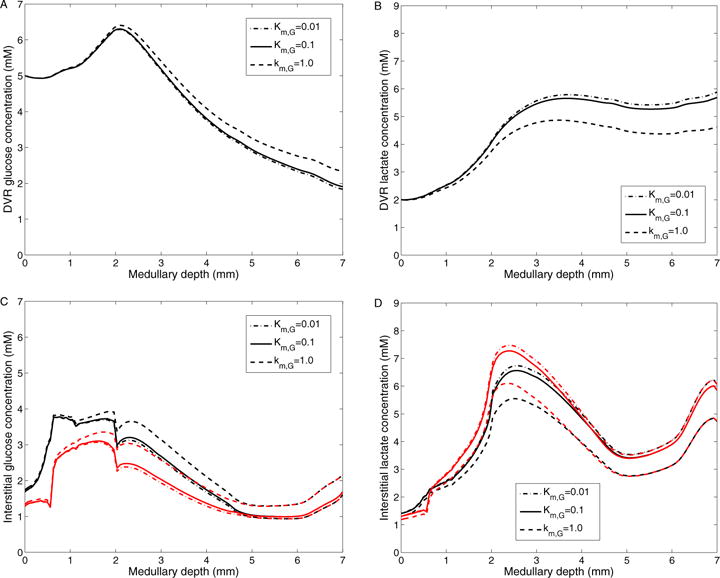
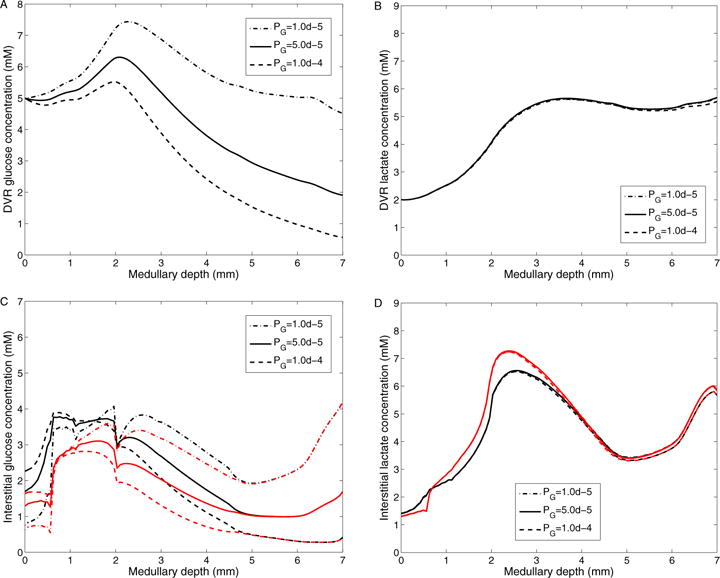
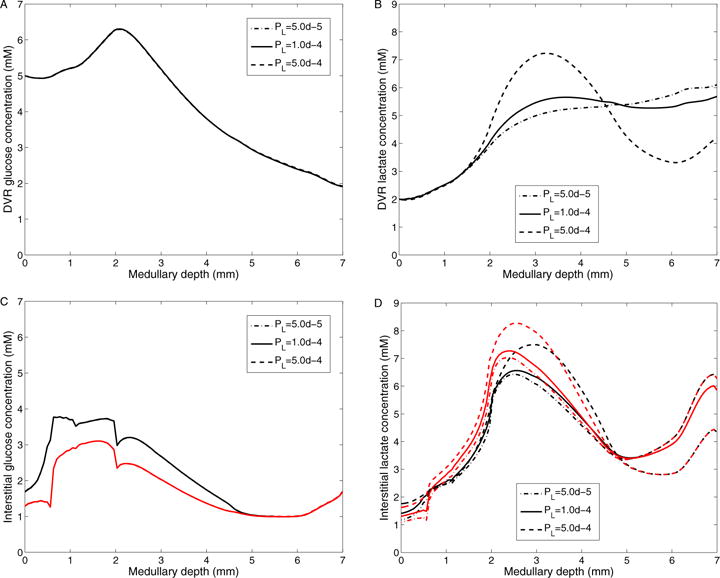
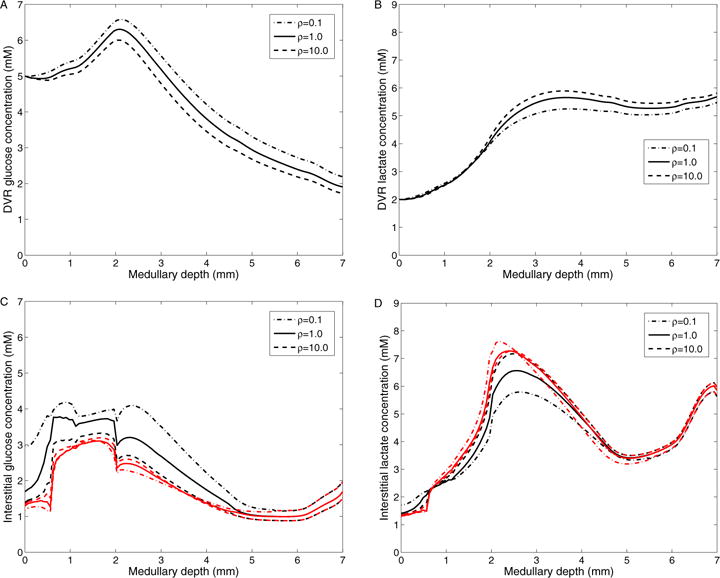
The metabolism of glucose provides most of the ATP required for energy-dependent transport processes. In the inner medulla of the mammalian kidney, limited blood flow and O2 supply yield low oxygen tension; therefore, a substantial fraction of the glucose metabolism in that region is anaerobic. Lactate is considered to be a waste product of anaerobic glycolysis, which yields two lactate molecules for each glucose molecule consumed, thereby likely leading to the production and accumulation of a significant amount of lactate in the inner medulla. To gain insights into the transport and metabolic processes in the kidney, we have developed a detailed mathematical model of the renal medulla of the rat kidney. The model represents the radial organization of the renal tubules and vessels, which centers around the vascular bundles in the outer medulla and around clusters of collecting ducts in the inner medulla. Model simulations yield significant radial gradients in interstitial fluid oxygen tension and glucose and lactate concentrations in the outer medulla and upper inner medulla. In the deep inner medulla, interstitial fluid concentrations become much more homogeneous, as the radial organization of tubules and vessels is not distinguishable. Using this model, we have identified parameters concerning glucose transport and basal metabolism, as well as lactate production via anaerobic glycolysis, that yield predicted blood glucose and lactate concentrations consistent with experimental measurements in the papillary tip. In addition, simulations indicate that the radial organization of the rat kidney may affect lactate buildup in the inner medulla.
Keywords: Anaerobic respiration; Glycolysis; Renal physiology.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Impacts of nitric oxide and superoxide on renal medullary oxygen transport and urine concentration.Am J Physiol Renal Physiol. 2015 May 1;308(9):F967-80. doi: 10.1152/ajprenal.00600.2014. Epub 2015 Jan 28. Am J Physiol Renal Physiol. 2015. PMID: 25651567 Free PMC article.
-
Modeling Glucose Metabolism in the Kidney.Bull Math Biol. 2016 Jun;78(6):1318-36. doi: 10.1007/s11538-016-0188-7. Epub 2016 Jul 1. Bull Math Biol. 2016. PMID: 27371260 Free PMC article.
-
Inner medullary lactate production and accumulation: a vasa recta model.Am J Physiol Renal Physiol. 2000 Sep;279(3):F468-81. doi: 10.1152/ajprenal.2000.279.3.F468. Am J Physiol Renal Physiol. 2000. PMID: 10966926
-
Is the function of the renal papilla coupled exclusively to an anaerobic pattern of metabolism?Am J Physiol. 1979 May;236(5):F423-33. doi: 10.1152/ajprenal.1979.236.5.F423. Am J Physiol. 1979. PMID: 220881 Review.
-
Recent advances in renal tubular biochemistry.Annu Rev Physiol. 1976;38:46-68. doi: 10.1146/annurev.ph.38.030176.000402. Annu Rev Physiol. 1976. PMID: 130828 Review. No abstract available.
Cited by
-
Relative Hypoxia and Early Diabetic Kidney Disease in Type 1 Diabetes.Diabetes. 2020 Dec;69(12):2700-2708. doi: 10.2337/db20-0457. Epub 2020 Jul 31. Diabetes. 2020. PMID: 32737116 Free PMC article.
-
Does Gadolinium Deposition Lead to Metabolite Alteration in the Dentate Nucleus? An MRS Study in Patients with MS.AJNR Am J Neuroradiol. 2022 Oct;43(10):1403-1410. doi: 10.3174/ajnr.A7623. Epub 2022 Sep 1. AJNR Am J Neuroradiol. 2022. PMID: 36574329 Free PMC article.
-
Youth versus adult-onset type 2 diabetic kidney disease: Insights into currently known structural differences and the potential underlying mechanisms.Clin Sci (Lond). 2022 Nov 11;136(21):1471-1483. doi: 10.1042/CS20210627. Clin Sci (Lond). 2022. PMID: 36326718 Free PMC article.
-
Mitochondrial Dysfunction and Diabetic Nephropathy: Nontraditional Therapeutic Opportunities.J Diabetes Res. 2021 Dec 9;2021:1010268. doi: 10.1155/2021/1010268. eCollection 2021. J Diabetes Res. 2021. PMID: 34926696 Free PMC article. Review.
-
Kidney physiology and susceptibility to acute kidney injury: implications for renoprotection.Nat Rev Nephrol. 2021 May;17(5):335-349. doi: 10.1038/s41581-021-00394-7. Epub 2021 Feb 5. Nat Rev Nephrol. 2021. PMID: 33547418 Review.
References
-
- Bagnasco S, Good D, Balaban R, Burg M. Lactate production in isolated segments of the rat nephron. Am J Physiol Renal Physiol. 1985;248:F522–F526. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
