

High-throughput and site-specific identification of 2'- O-methylation sites using ribose oxidation sequencing (RibOxi-seq)
- PMID: 28495677
- PMCID: PMC5513074
- DOI: 10.1261/rna.061549.117
High-throughput and site-specific identification of 2'- O-methylation sites using ribose oxidation sequencing (RibOxi-seq)
Abstract
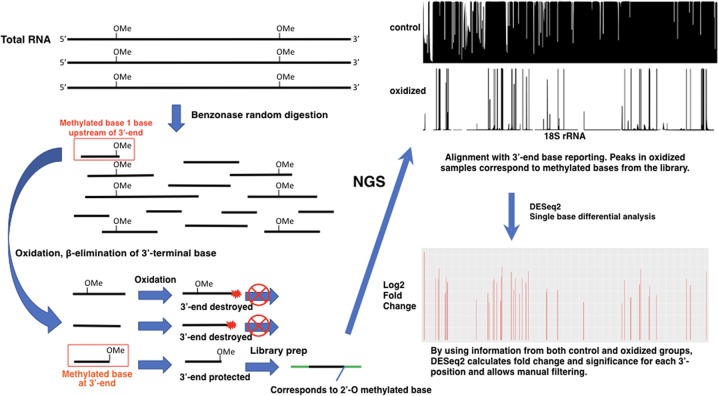
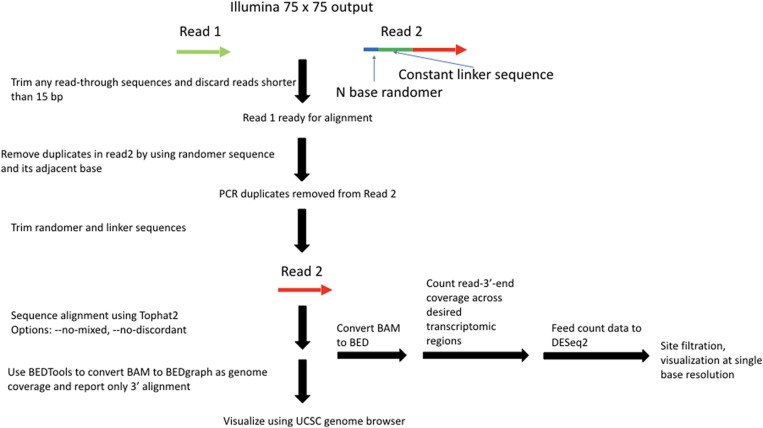
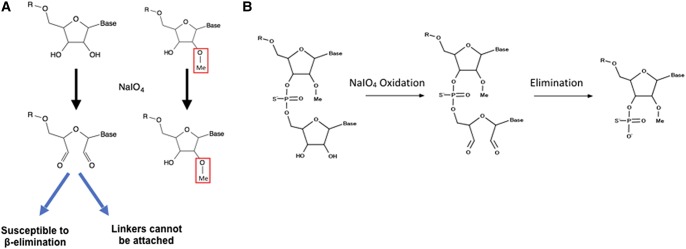
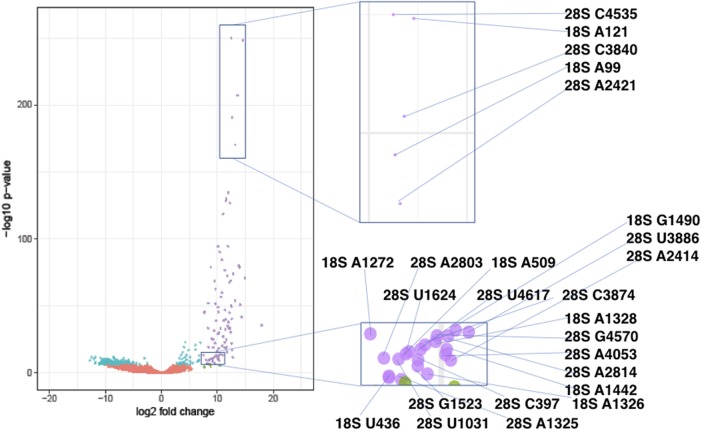
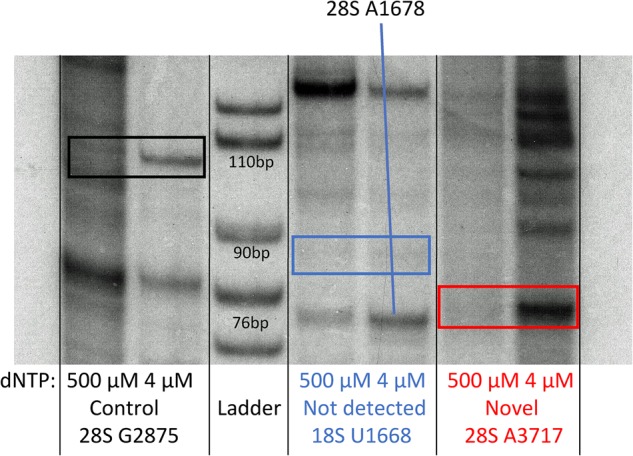
Ribose methylation (2'-O-methylation, 2'-OMe) occurs at high frequencies in rRNAs and other small RNAs and is carried out using a shared mechanism across eukaryotes and archaea. As RNA modifications are important for ribosome maturation, and alterations in these modifications are associated with cellular defects and diseases, it is important to characterize the landscape of 2'-O-methylation. Here we report the development of a highly sensitive and accurate method for ribose methylation detection using next-generation sequencing. A key feature of this method is the generation of RNA fragments with random 3'-ends, followed by periodate oxidation of all molecules terminating in 2',3'-OH groups. This allows only RNAs harboring 2'-OMe groups at their 3'-ends to be sequenced. Although currently requiring microgram amounts of starting material, this method is robust for the analysis of rRNAs even at low sequencing depth.
Keywords: 2′-O-methylation; RNA editing; RNA modification; ribosomal RNA.
© 2017 Zhu et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Decatur WA, Fournier MJ. 2002. rRNA modifications and ribosome function. Trends Biochem Sci 27: 344–351. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials