

The uniqueness of subunit α of mycobacterial F-ATP synthases: An evolutionary variant for niche adaptation
- PMID: 28495884
- PMCID: PMC5500794
- DOI: 10.1074/jbc.M117.784959
The uniqueness of subunit α of mycobacterial F-ATP synthases: An evolutionary variant for niche adaptation
Abstract
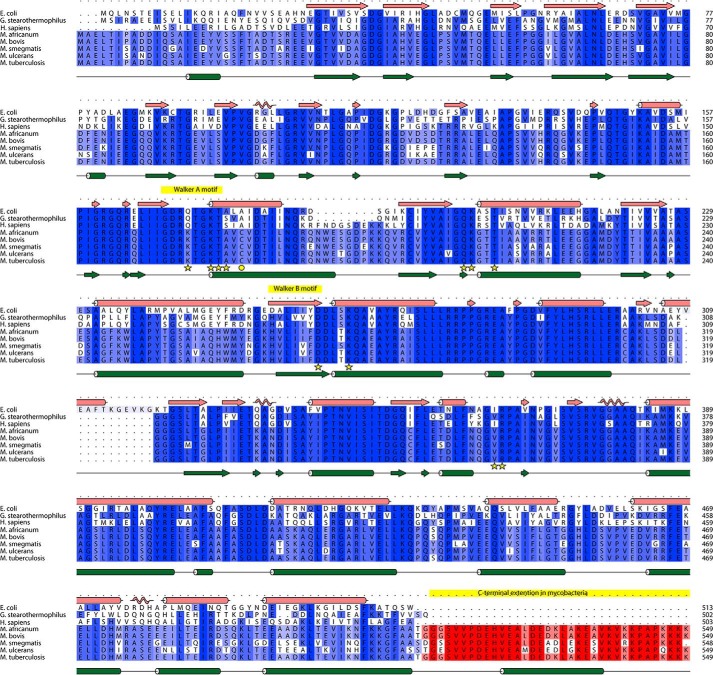
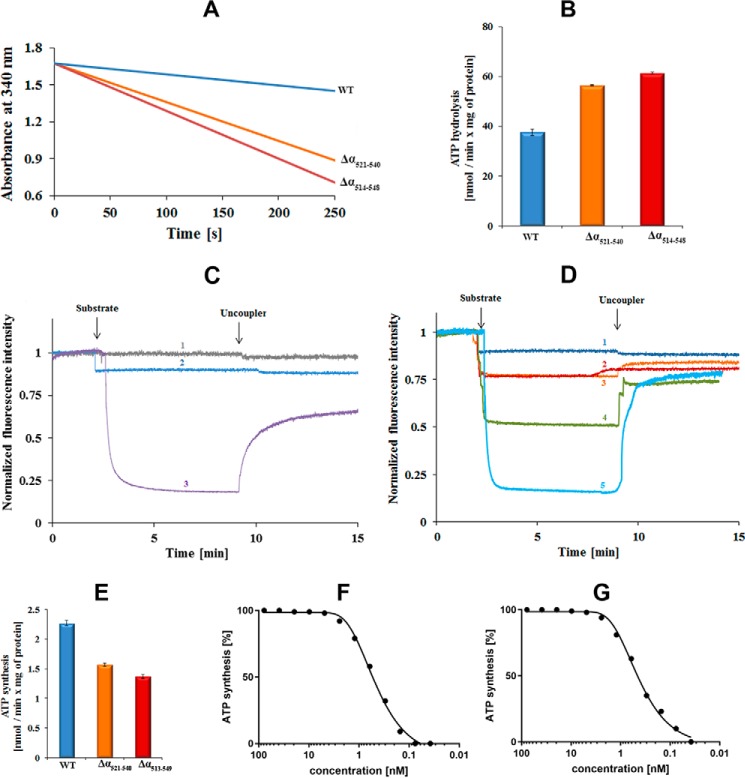
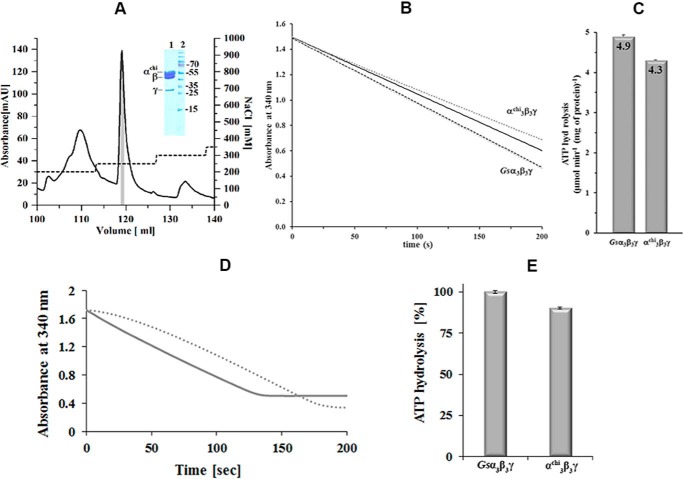
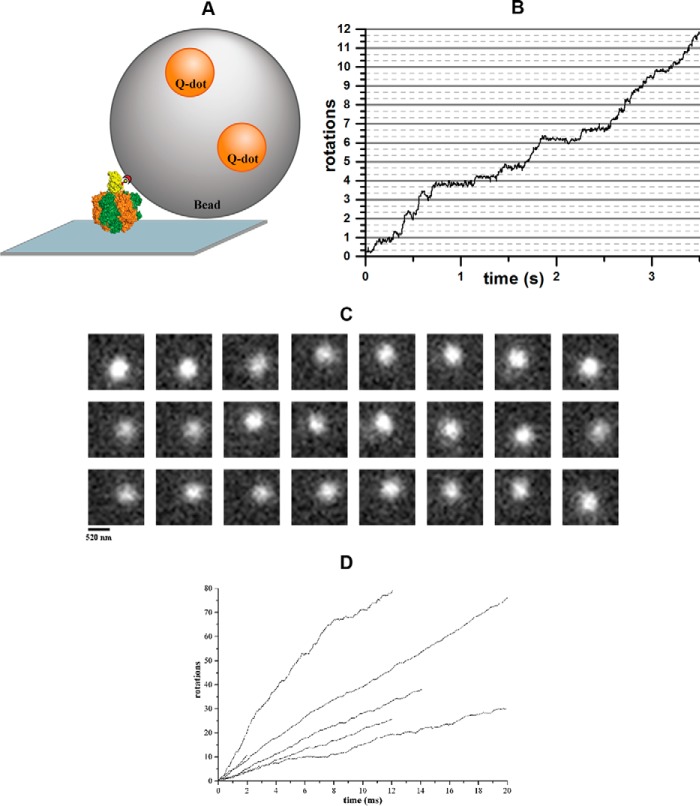
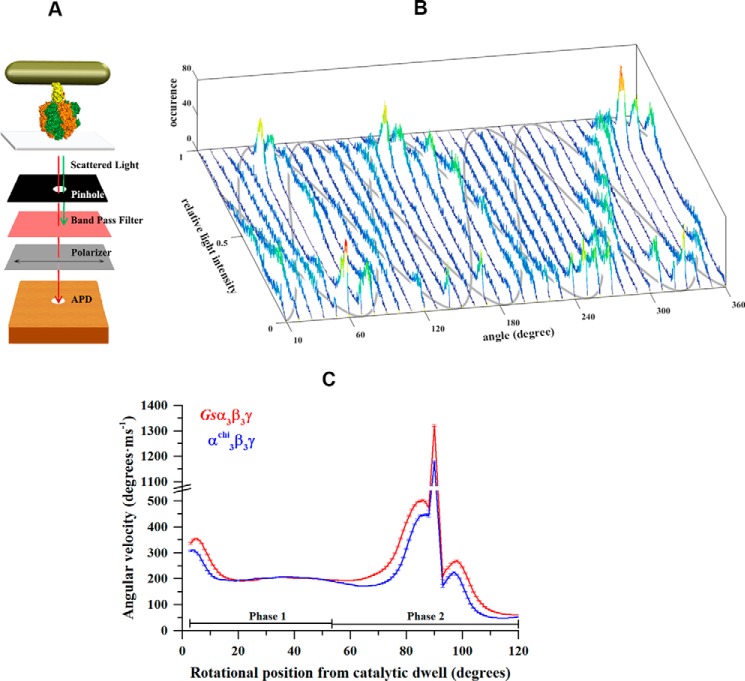
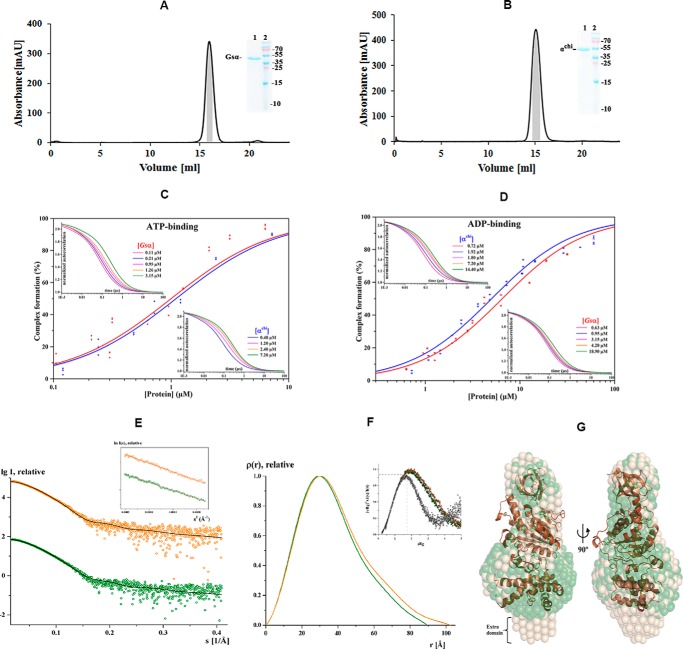
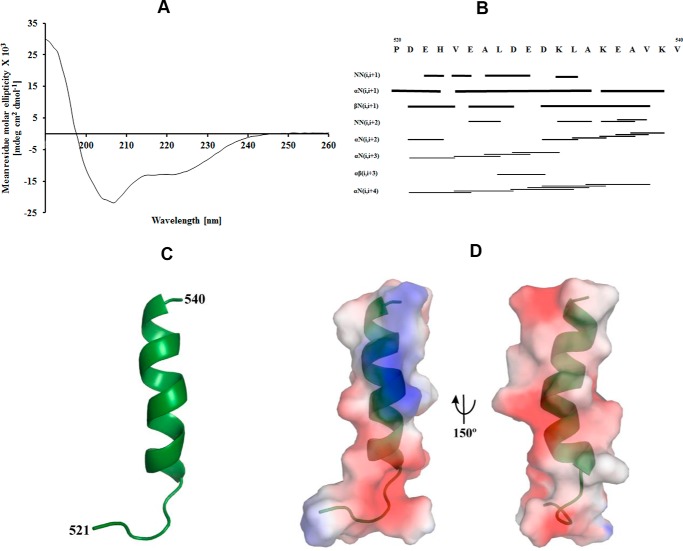
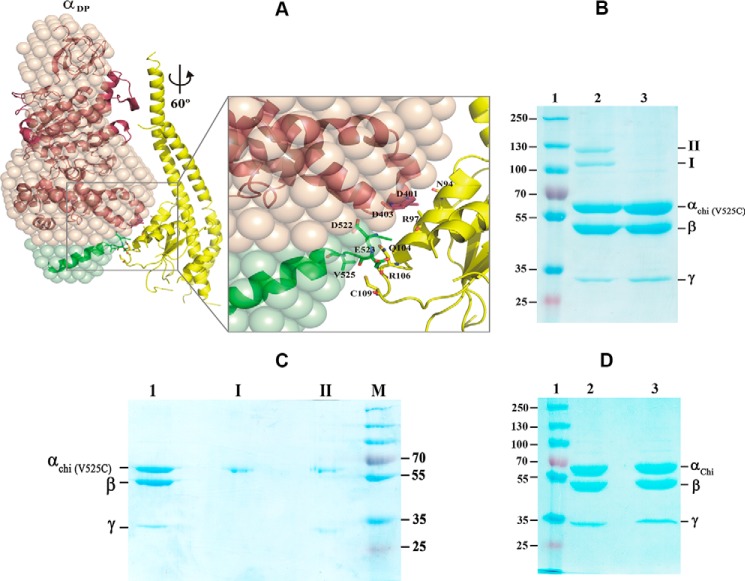
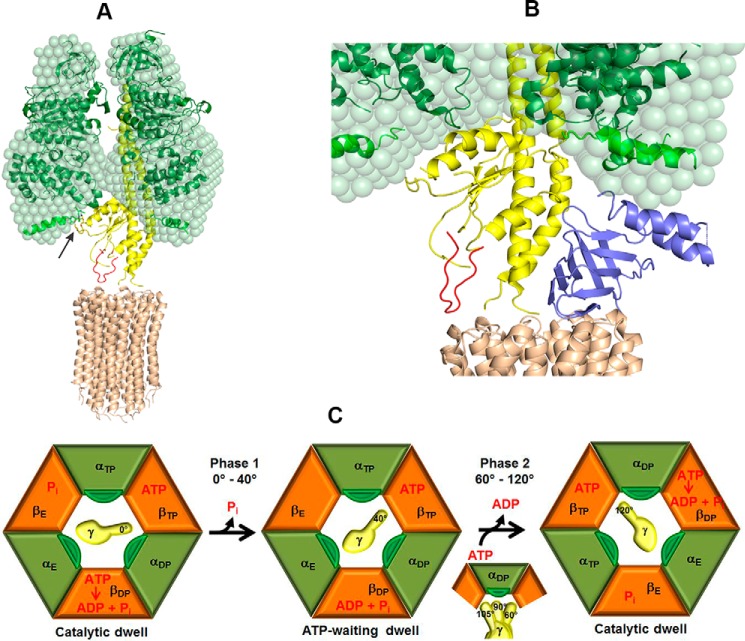
The F1F0 -ATP (F-ATP) synthase is essential for growth of Mycobacterium tuberculosis, the causative agent of tuberculosis (TB). In addition to their synthase function most F-ATP synthases possess an ATP-hydrolase activity, which is coupled to proton-pumping activity. However, the mycobacterial enzyme lacks this reverse activity, but the reason for this deficiency is unclear. Here, we report that a Mycobacterium-specific, 36-amino acid long C-terminal domain in the nucleotide-binding subunit α (Mtα) of F-ATP synthase suppresses its ATPase activity and determined the mechanism of suppression. First, we employed vesicles to show that in intact membrane-embedded mycobacterial F-ATP synthases deletion of the C-terminal domain enabled ATPase and proton-pumping activity. We then generated a heterologous F-ATP synthase model system, which demonstrated that transfer of the mycobacterial C-terminal domain to a standard F-ATP synthase α subunit suppresses ATPase activity. Single-molecule rotation assays indicated that the introduction of this Mycobacterium-specific domain decreased the angular velocity of the power-stroke after ATP binding. Solution X-ray scattering data and NMR results revealed the solution shape of Mtα and the 3D structure of the subunit α C-terminal peptide 521PDEHVEALDEDKLAKEAVKV540 of M. tubercolosis (Mtα(521-540)), respectively. Together with cross-linking studies, the solution structural data lead to a model, in which Mtα(521-540) comes in close proximity with subunit γ residues 104-109, whose interaction may influence the rotation of the camshaft-like subunit γ. Finally, we propose that the unique segment Mtα(514-549), which is accessible at the C terminus of mycobacterial subunit α, is a promising drug epitope.
Keywords: ATP synthase; F-ATP synthase; F1FO-ATPase; Mycobacterium; bioenergetics; membrane protein; subunit α; tuberculosis.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Sassetti C. M., Boyd D. H., and Rubin E. J. (2003) Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 48, 77–84 - PubMed
-
- Lu P., Lill H., and Bald D. (2014) ATP synthase in mycobacteria: special features and implications for a function as drug target. Biochim. Biophys. Acta 1837, 1208–1218 - PubMed
-
- Cox R. A., and Cook G. M. (2007) Growth regulation in the mycobacterial cell. Curr. Mol. Med. 7, 231–245 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
