

An Evolutionarily Conserved Role of Presenilin in Neuronal Protection in the Aging Drosophila Brain
- PMID: 28495961
- PMCID: PMC5500145
- DOI: 10.1534/genetics.116.196881
An Evolutionarily Conserved Role of Presenilin in Neuronal Protection in the Aging Drosophila Brain
Abstract
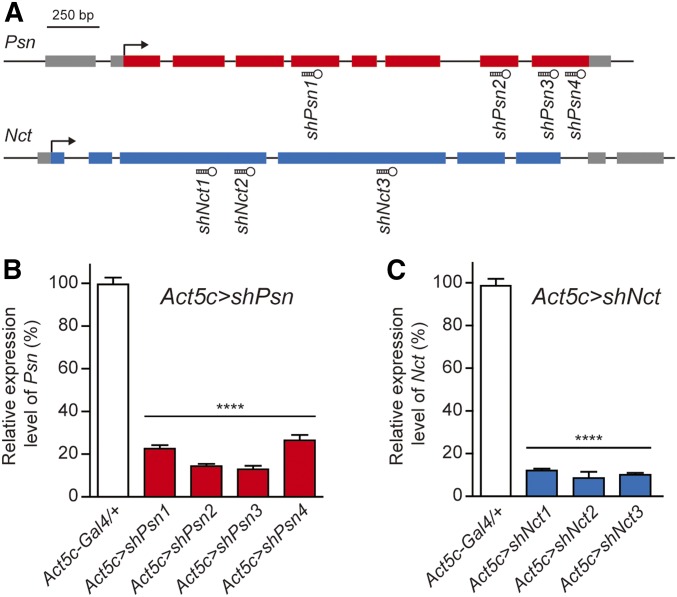
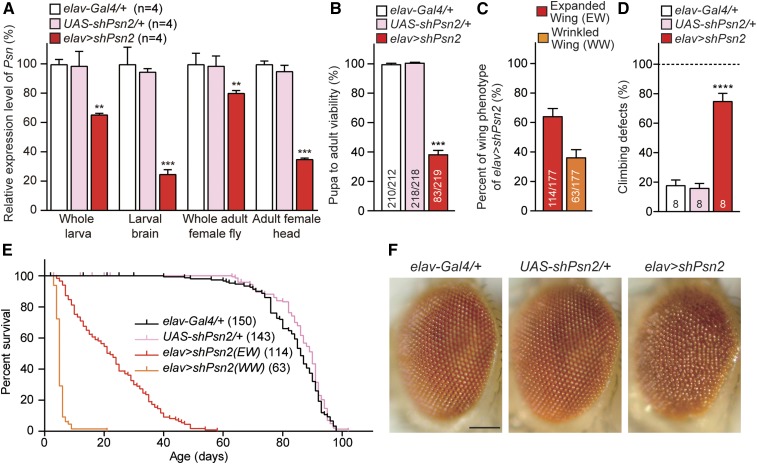
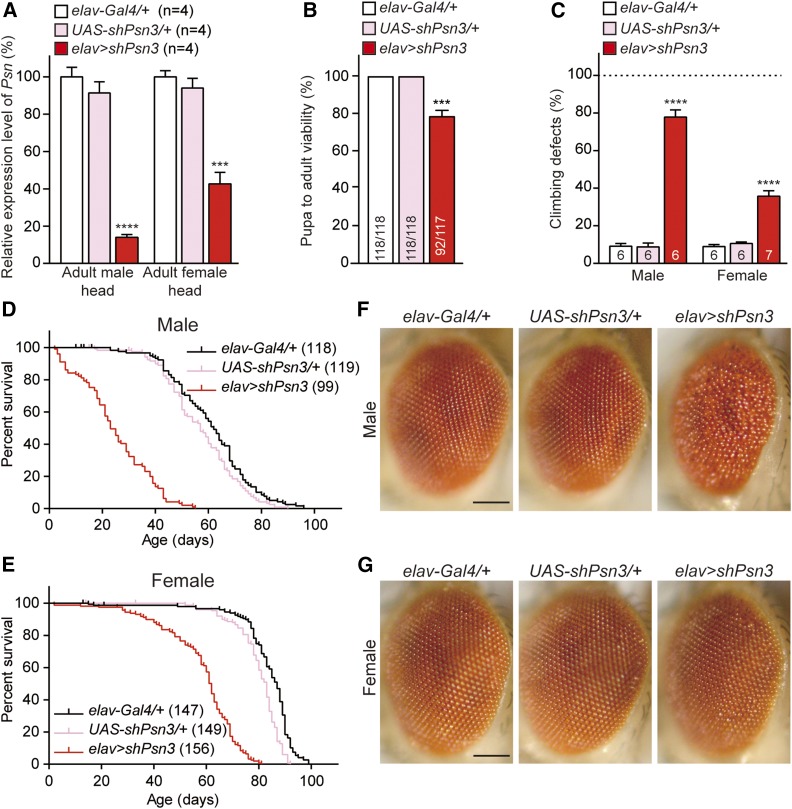
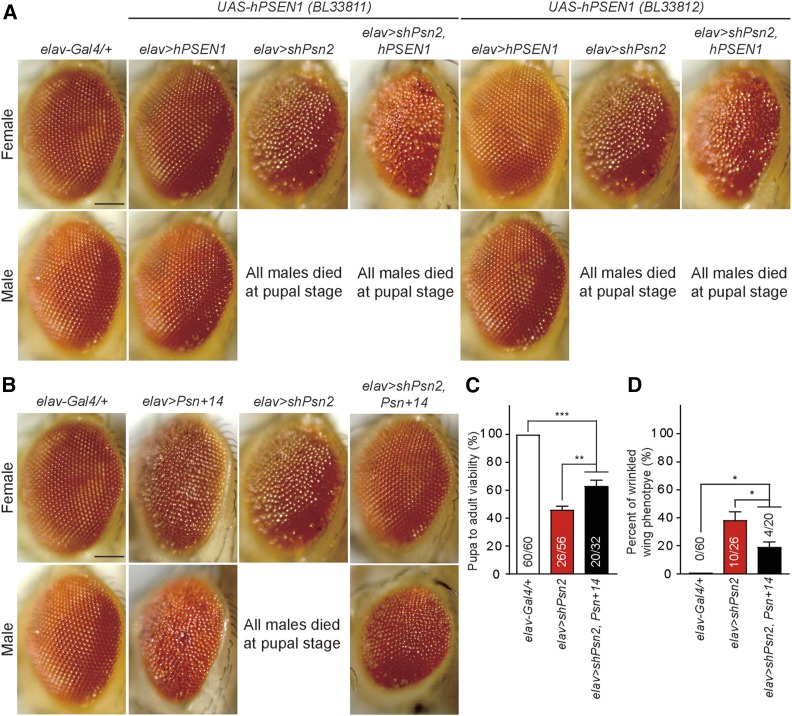
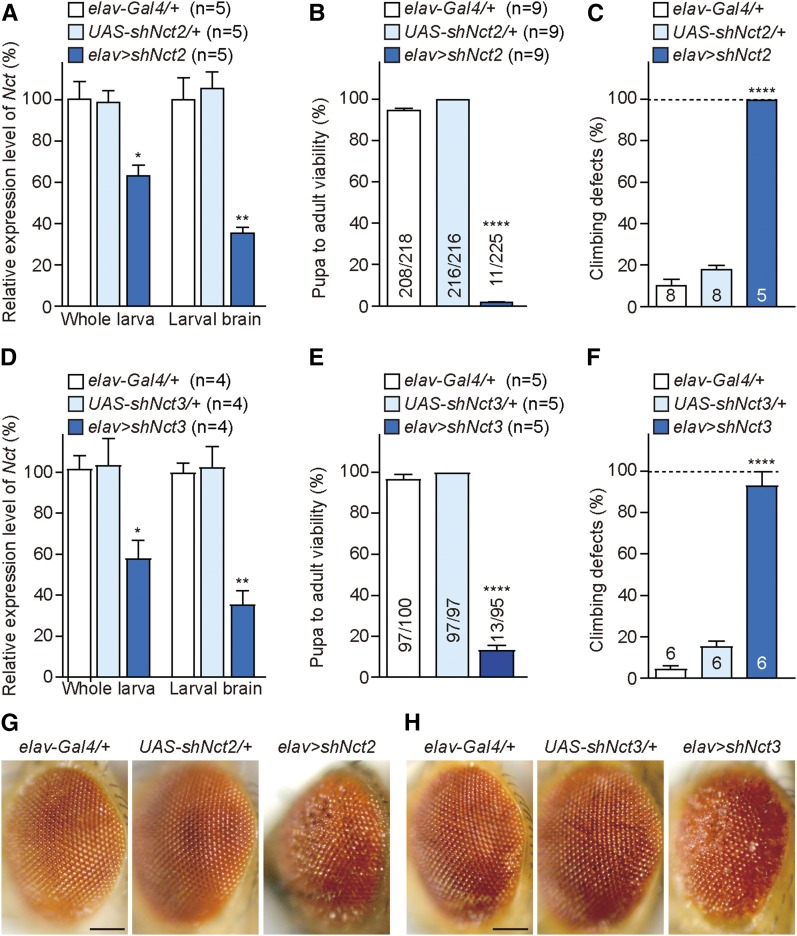
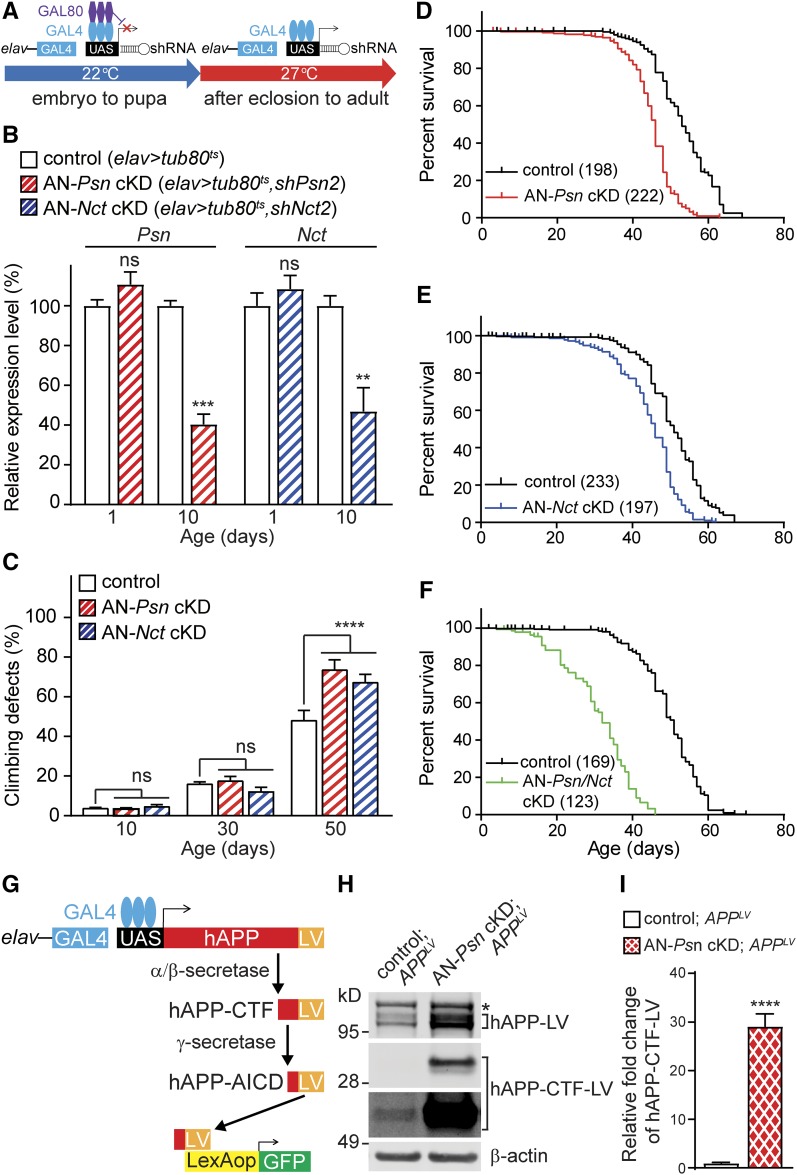
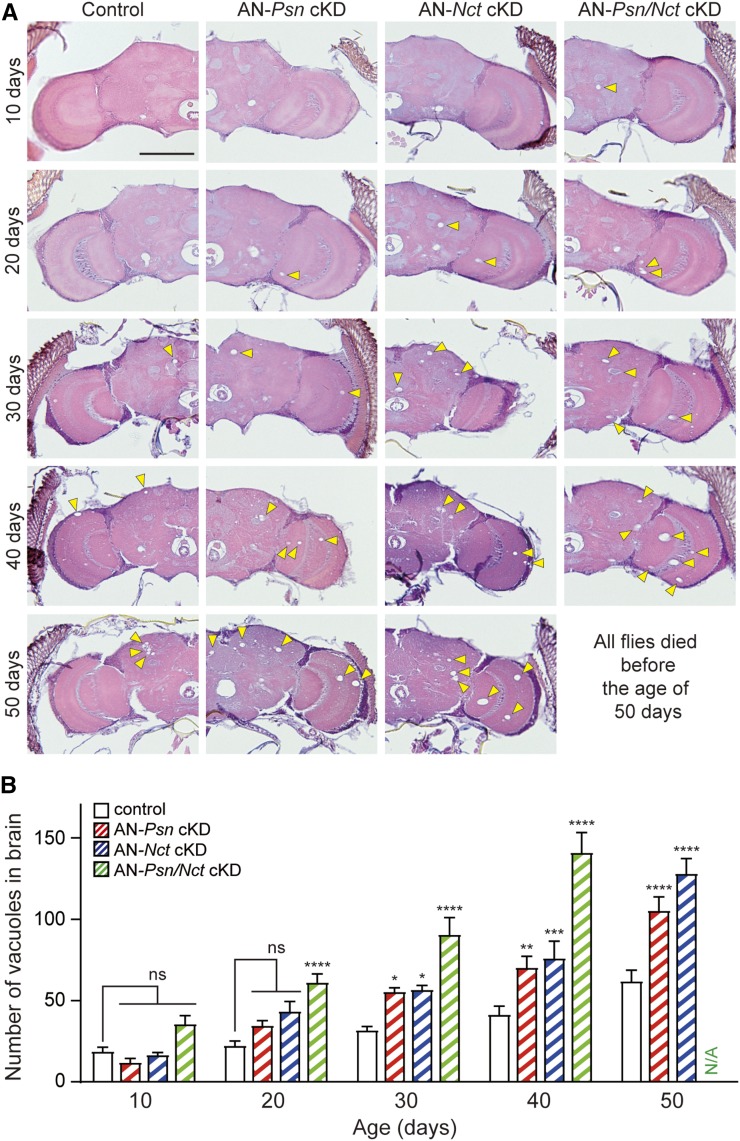
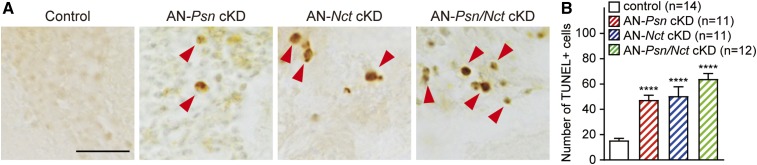
Mutations in the Presenilin genes are the major genetic cause of Alzheimer's disease. Presenilin and Nicastrin are essential components of γ-secretase, a multi-subunit protease that cleaves Type I transmembrane proteins. Genetic studies in mice previously demonstrated that conditional inactivation of Presenilin or Nicastrin in excitatory neurons of the postnatal forebrain results in memory deficits, synaptic impairment, and age-dependent neurodegeneration. The roles of Drosophila Presenilin (Psn) and Nicastrin (Nct) in the adult fly brain, however, are unknown. To knockdown (KD) Psn or Nct selectively in neurons of the adult brain, we generated multiple shRNA lines. Using a ubiquitous driver, these shRNA lines resulted in 80-90% reduction of mRNA and pupal lethality-a phenotype that is shared with Psn and Nct mutants carrying nonsense mutations. Furthermore, expression of these shRNAs in the wing disc caused notching wing phenotypes, which are also shared with Psn and Nct mutants. Similar to Nct, neuron-specific Psn KD using two independent shRNA lines led to early mortality and rough eye phenotypes, which were rescued by a fly Psn transgene. Interestingly, conditional KD (cKD) of Psn or Nct in adult neurons using the elav-Gal4 and tubulin-Gal80ts system caused shortened lifespan, climbing defects, increases in apoptosis, and age-dependent neurodegeneration. Together, these findings demonstrate that, similar to their mammalian counterparts, Drosophila Psn and Nct are required for neuronal survival during aging and normal lifespan, highlighting an evolutionarily conserved role of Presenilin in neuronal protection in the aging brain.
Keywords: Alzheimer’s disease; brain; conditional knockdown; shRNA; γ-secretase.
Copyright © 2017 by the Genetics Society of America.
Figures

Similar articles
-
Lipophorin receptors genetically modulate neurodegeneration caused by reduction of Psn expression in the aging Drosophila brain.Genetics. 2024 Jan 3;226(1):iyad202. doi: 10.1093/genetics/iyad202. Genetics. 2024. PMID: 37996068 Free PMC article.
-
Aph-1 is required to regulate Presenilin-mediated gamma-secretase activity and cell survival in Drosophila wing development.Genesis. 2009 Mar;47(3):169-74. doi: 10.1002/dvg.20478. Genesis. 2009. PMID: 19191330 Free PMC article.
-
Synaptic function of nicastrin in hippocampal neurons.Proc Natl Acad Sci U S A. 2014 Jun 17;111(24):8973-8. doi: 10.1073/pnas.1408554111. Epub 2014 Jun 2. Proc Natl Acad Sci U S A. 2014. PMID: 24889619 Free PMC article.
-
[Advances on Drosophila presenilin gene].Yi Chuan. 2011 Nov;33(11):1164-70. doi: 10.3724/sp.j.1005.2011.01164. Yi Chuan. 2011. PMID: 22120070 Review. Chinese.
-
Contribution of the Presenilins in the cell biology, structure and function of γ-secretase.Semin Cell Dev Biol. 2020 Sep;105:12-26. doi: 10.1016/j.semcdb.2020.02.005. Epub 2020 Mar 4. Semin Cell Dev Biol. 2020. PMID: 32146031 Review.
Cited by
-
Regulation of autophagy, lipid metabolism, and neurodegenerative pathology by heparan sulfate proteoglycans.Front Genet. 2023 Jan 9;13:1012706. doi: 10.3389/fgene.2022.1012706. eCollection 2022. Front Genet. 2023. PMID: 36699460 Free PMC article.
-
Glycogen synthase kinase 3β (GSK3β) and presenilin (PS) are key regulators of kinesin-1-mediated cargo motility within axons.Front Cell Dev Biol. 2023 Jun 9;11:1202307. doi: 10.3389/fcell.2023.1202307. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37363727 Free PMC article. Review.
-
Identification of presenilin mutations that have sufficient gamma-secretase proteolytic activity to mediate Notch signaling but disrupt organelle and neuronal health.Neurobiol Dis. 2025 Aug;212:106961. doi: 10.1016/j.nbd.2025.106961. Epub 2025 May 20. Neurobiol Dis. 2025. PMID: 40404063 Free PMC article.
-
Ensheathing glia promote increased lifespan and healthy brain aging.Aging Cell. 2023 May;22(5):e13803. doi: 10.1111/acel.13803. Epub 2023 Feb 24. Aging Cell. 2023. PMID: 36840361 Free PMC article.
-
TM2D genes regulate Notch signaling and neuronal function in Drosophila.PLoS Genet. 2021 Dec 14;17(12):e1009962. doi: 10.1371/journal.pgen.1009962. eCollection 2021 Dec. PLoS Genet. 2021. PMID: 34905536 Free PMC article.
References
-
- Beglopoulos V., Sun X., Saura C. A., Lemere C. A., Kim R. D., et al. , 2004. Reduced β-amyloid production and increased inflammatory responses in presenilin conditional knock-out mice. J. Biol. Chem. 279: 46907–46914. - PubMed
-
- Brand A. H., Perrimon N., 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
