

Potent antitumour activity of interleukin-2-Fc fusion proteins requires Fc-mediated depletion of regulatory T-cells
- PMID: 28497796
- PMCID: PMC5437307
- DOI: 10.1038/ncomms15373
Potent antitumour activity of interleukin-2-Fc fusion proteins requires Fc-mediated depletion of regulatory T-cells
Abstract
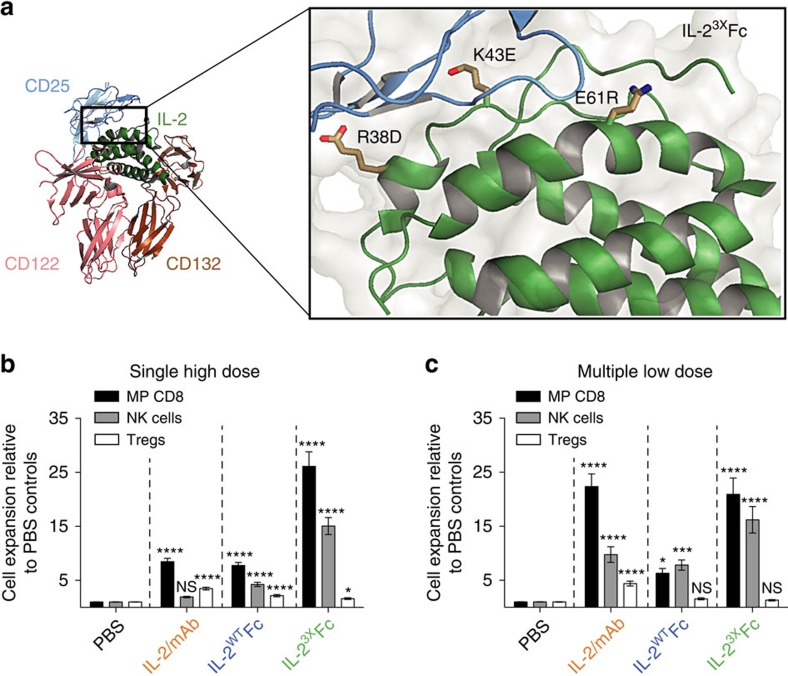
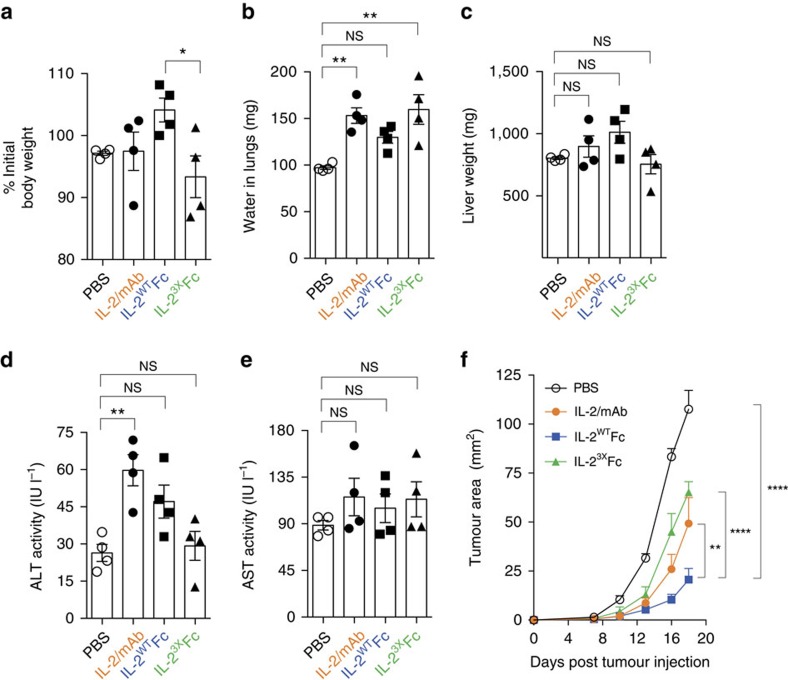
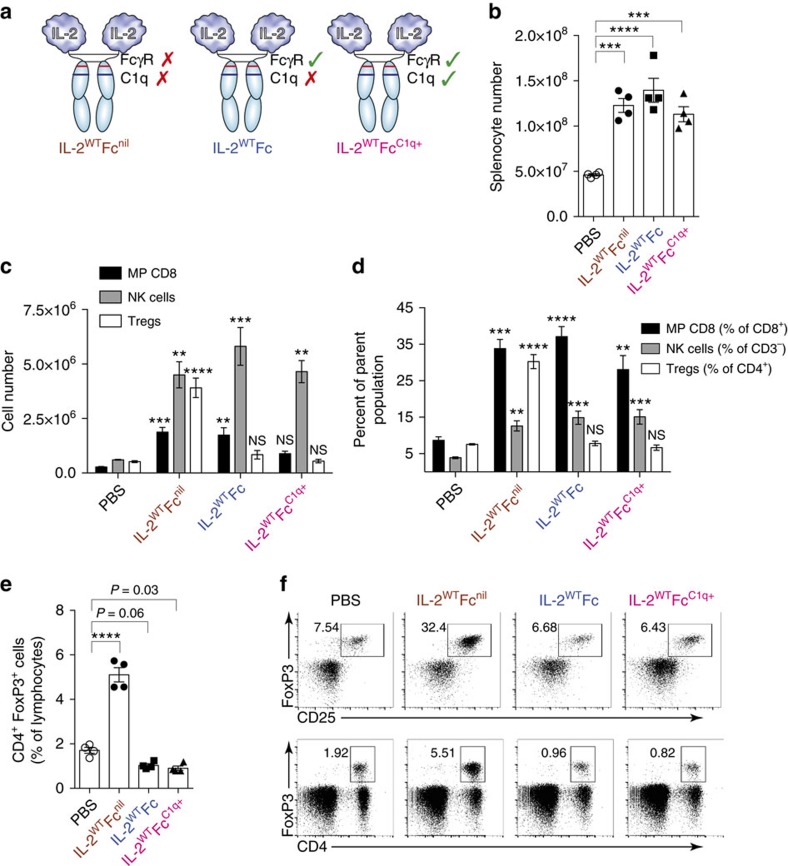
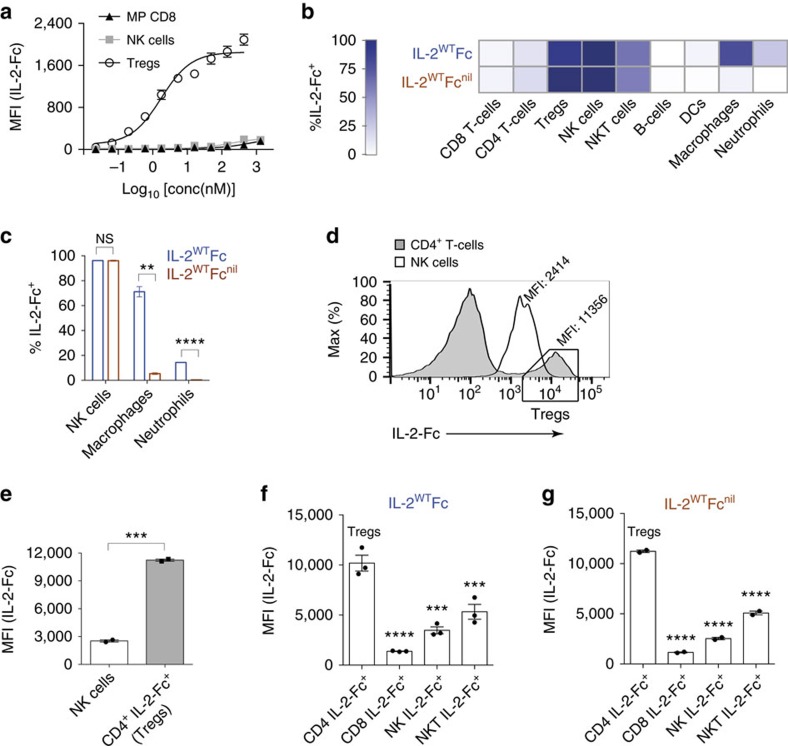
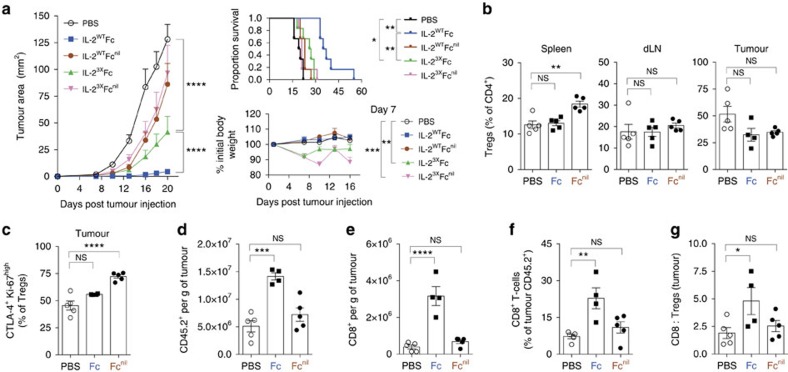
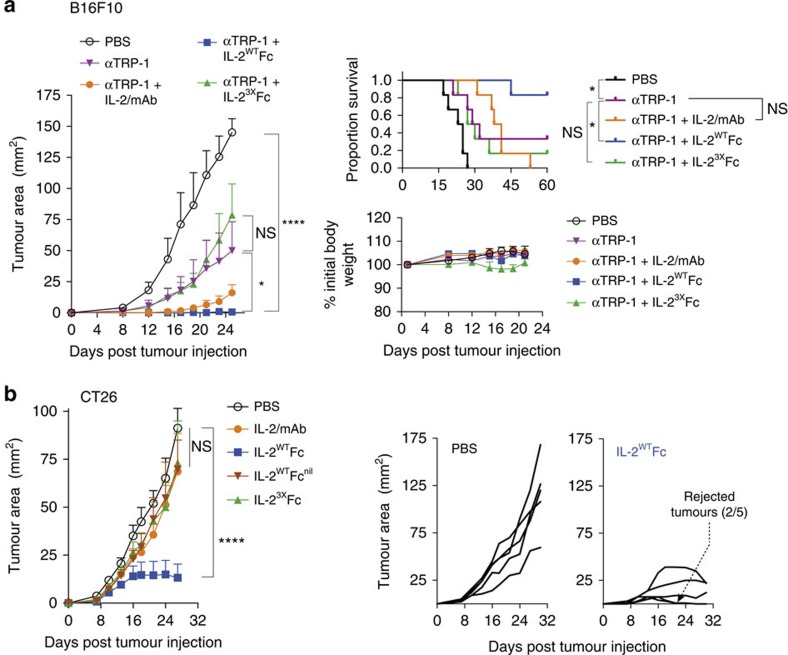
Interleukin-2 (IL-2) is an established therapeutic agent used for cancer immunotherapy. Since treatment efficacy is mediated by CD8+ and NK cell activity at the tumour site, considerable efforts have focused on generating variants that expand these subsets systemically, as exemplified by IL-2/antibody complexes and 'superkines'. Here we describe a novel determinant of antitumour activity using fusion proteins consisting of IL-2 and the antibody fragment crystallizable (Fc) region. Generation of long-lived IL-2-Fc variants in which CD25 binding is abolished through mutation effectively prevents unwanted activation of CD25+ regulatory T-cells (Tregs) and results in strong expansion of CD25- cytotoxic subsets. Surprisingly, however, such variants are less effective than wild-type IL-2-Fc in mediating tumour rejection. Instead, we report that efficacy is crucially dependent on depletion of Tregs through Fc-mediated immune effector functions. Our results underpin an unexpected mechanism of action and provide important guidance for the development of next generation IL-2 therapeutics.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
ALKS 4230: a novel engineered IL-2 fusion protein with an improved cellular selectivity profile for cancer immunotherapy.J Immunother Cancer. 2020 Apr;8(1):e000673. doi: 10.1136/jitc-2020-000673. J Immunother Cancer. 2020. PMID: 32317293 Free PMC article.
-
Improved cancer immunotherapy by a CD25-mimobody conferring selectivity to human interleukin-2.Sci Transl Med. 2016 Nov 30;8(367):367ra166. doi: 10.1126/scitranslmed.aag3187. Sci Transl Med. 2016. PMID: 27903862
-
IL-15R alpha-IgG1-Fc enhances IL-2 and IL-15 anti-tumor action through NK and CD8+ T cells proliferation and activation.J Mol Cell Biol. 2010 Aug;2(4):217-22. doi: 10.1093/jmcb/mjq012. J Mol Cell Biol. 2010. PMID: 20671116
-
Increasing the biological activity of IL-2 and IL-15 through complexing with anti-IL-2 mAbs and IL-15Rα-Fc chimera.Immunol Lett. 2014 May-Jun;159(1-2):1-10. doi: 10.1016/j.imlet.2014.01.017. Epub 2014 Feb 7. Immunol Lett. 2014. PMID: 24512738 Review.
-
Selectively expanding subsets of T cells in mice by injection of interleukin-2/antibody complexes: implications for transplantation tolerance.Transplant Proc. 2012 May;44(4):1032-4. doi: 10.1016/j.transproceed.2012.01.093. Transplant Proc. 2012. PMID: 22564618 Review.
Cited by
-
Intratumoral nanobody-IL-2 fusions that bind the tumor extracellular matrix suppress solid tumor growth in mice.PNAS Nexus. 2022 Nov 3;1(5):pgac244. doi: 10.1093/pnasnexus/pgac244. eCollection 2022 Nov. PNAS Nexus. 2022. PMID: 36712341 Free PMC article.
-
The use of supercytokines, immunocytokines, engager cytokines, and other synthetic cytokines in immunotherapy.Cell Mol Immunol. 2022 Feb;19(2):192-209. doi: 10.1038/s41423-021-00786-6. Epub 2022 Jan 4. Cell Mol Immunol. 2022. PMID: 35043005 Free PMC article. Review.
-
Human interleukin-2 receptor β mutations associated with defects in immunity and peripheral tolerance.J Exp Med. 2019 Jun 3;216(6):1311-1327. doi: 10.1084/jem.20182304. Epub 2019 Apr 30. J Exp Med. 2019. PMID: 31040185 Free PMC article.
-
Molecular reshaping of phage-displayed Interleukin-2 at beta chain receptor interface to obtain potent super-agonists with improved developability profiles.Commun Biol. 2023 Aug 9;6(1):828. doi: 10.1038/s42003-023-05188-0. Commun Biol. 2023. PMID: 37558752 Free PMC article.
-
Pharmacokinetics, pharmacodynamics, and toxicity of a PD-1-targeted IL-15 in cynomolgus monkeys.PLoS One. 2024 Feb 5;19(2):e0298240. doi: 10.1371/journal.pone.0298240. eCollection 2024. PLoS One. 2024. PMID: 38315680 Free PMC article.
References
-
- Boyman O. & Sprent J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat. Rev. Immunol. 12, 180–190 (2012). - PubMed
-
- Morgan D. A., Ruscetti F. W. & Gallo R. Selective in vitro growth of T-lymphocytes from normal human bone marrows. Science 193, 1007–1008 (1976). - PubMed
-
- Taniguchi T. et al.. Structure and expression of a cloned cDNA for human interleukin-2. Nature 302, 305–310 (1983). - PubMed
-
- Lotze M. T. et al.. High-dose recombinant interleukin 2 in the treatment of patients with disseminated cancer: responses, treatment-related morbidity, and histologic findings. JAMA 256, 3117–3124 (1986). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
