

A viral suppressor of RNA silencing inhibits ARGONAUTE 1 function by precluding target RNA binding to pre-assembled RISC
- PMID: 28499009
- PMCID: PMC5737661
- DOI: 10.1093/nar/gkx379
A viral suppressor of RNA silencing inhibits ARGONAUTE 1 function by precluding target RNA binding to pre-assembled RISC
Abstract
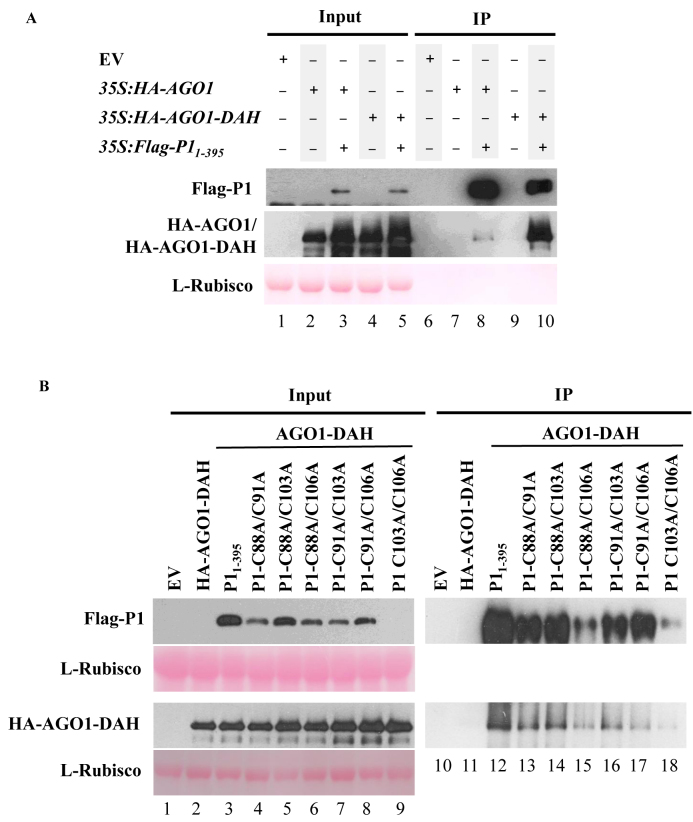
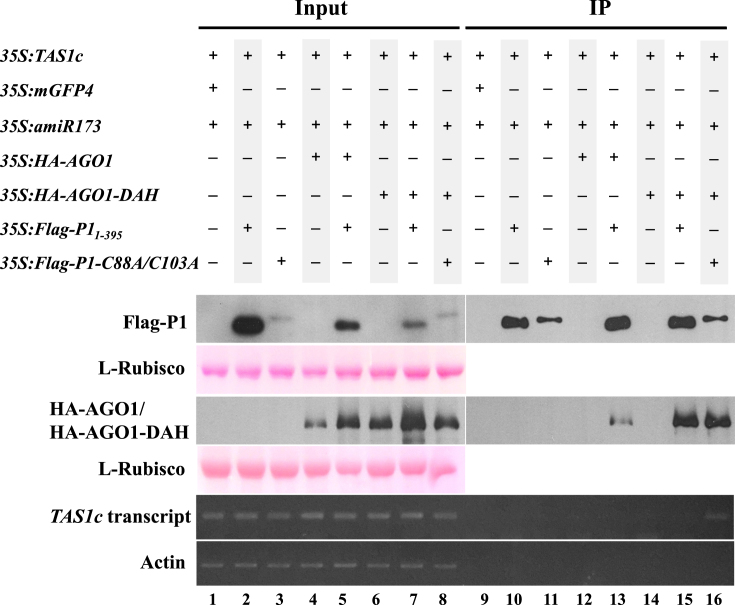
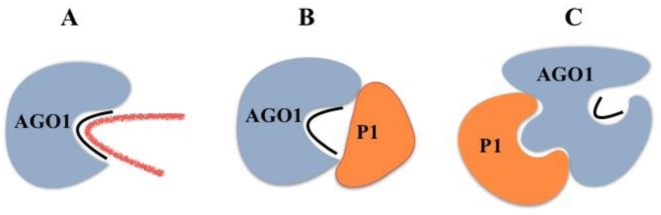
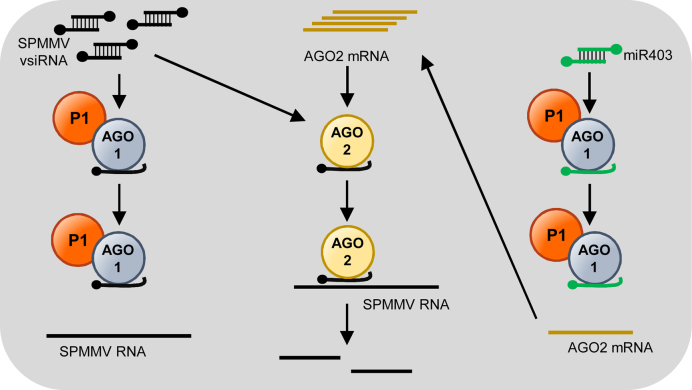
In most eukaryotes, RNA silencing is an adaptive immune system regulating key biological processes including antiviral defense. To evade this response, viruses of plants, worms and insects have evolved viral suppressors of RNA silencing proteins (VSRs). Various VSRs, such as P1 from Sweet potato mild mottle virus (SPMMV), inhibit the activity of RNA-induced silencing complexes (RISCs) including an ARGONAUTE (AGO) protein loaded with a small RNA. However, the specific mechanisms explaining this class of inhibition are unknown. Here, we show that SPMMV P1 interacts with AGO1 and AGO2 from Arabidopsis thaliana, but solely interferes with AGO1 function. Moreover, a mutational analysis of a newly identified zinc finger domain in P1 revealed that this domain could represent an effector domain as it is required for P1 suppressor activity but not for AGO1 binding. Finally, a comparative analysis of the target RNA binding capacity of AGO1 in the presence of wild-type or suppressor-defective P1 forms revealed that P1 blocks target RNA binding to AGO1. Our results describe the negative regulation of RISC, the small RNA containing molecular machine.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Viral protein inhibits RISC activity by argonaute binding through conserved WG/GW motifs.PLoS Pathog. 2010 Jul 15;6(7):e1000996. doi: 10.1371/journal.ppat.1000996. PLoS Pathog. 2010. PMID: 20657820 Free PMC article.
-
Degradation of the antiviral component ARGONAUTE1 by the autophagy pathway.Proc Natl Acad Sci U S A. 2012 Sep 25;109(39):15942-6. doi: 10.1073/pnas.1209487109. Epub 2012 Sep 10. Proc Natl Acad Sci U S A. 2012. PMID: 23019378 Free PMC article.
-
Cucumber mosaic virus-encoded 2b suppressor inhibits Arabidopsis Argonaute1 cleavage activity to counter plant defense.Genes Dev. 2006 Dec 1;20(23):3255-68. doi: 10.1101/gad.1495506. Genes Dev. 2006. PMID: 17158744 Free PMC article.
-
Structural insights into the arms race between host and virus along RNA silencing pathways in Arabidopsis thaliana.Biol Rev Camb Philos Soc. 2014 May;89(2):337-55. doi: 10.1111/brv.12057. Epub 2013 Sep 4. Biol Rev Camb Philos Soc. 2014. PMID: 24034134 Review.
-
Anatomy of RISC: how do small RNAs and chaperones activate Argonaute proteins?Wiley Interdiscip Rev RNA. 2016 Sep;7(5):637-60. doi: 10.1002/wrna.1356. Epub 2016 May 16. Wiley Interdiscip Rev RNA. 2016. PMID: 27184117 Free PMC article. Review.
Cited by
-
Antiviral Roles of Abscisic Acid in Plants.Front Plant Sci. 2017 Oct 11;8:1760. doi: 10.3389/fpls.2017.01760. eCollection 2017. Front Plant Sci. 2017. PMID: 29075279 Free PMC article. Review.
-
Proteome expansion in the Potyviridae evolutionary radiation.FEMS Microbiol Rev. 2022 Jul 1;46(4):fuac011. doi: 10.1093/femsre/fuac011. FEMS Microbiol Rev. 2022. PMID: 35195244 Free PMC article. Review.
-
Antiviral ARGONAUTEs Against Turnip Crinkle Virus Revealed by Image-Based Trait Analysis.Plant Physiol. 2019 Jul;180(3):1418-1435. doi: 10.1104/pp.19.00121. Epub 2019 May 1. Plant Physiol. 2019. PMID: 31043494 Free PMC article.
-
Generation of virus-resistant potato plants by RNA genome targeting.Plant Biotechnol J. 2019 Sep;17(9):1814-1822. doi: 10.1111/pbi.13102. Epub 2019 Mar 8. Plant Biotechnol J. 2019. PMID: 30803101 Free PMC article.
-
Assessment of the RNA Silencing Suppressor Activity of Protein P0 of Pepper Vein Yellows Virus 5: Uncovering Natural Variability, Relevant Motifs and Underlying Mechanism.Biology (Basel). 2022 Dec 12;11(12):1801. doi: 10.3390/biology11121801. Biology (Basel). 2022. PMID: 36552310 Free PMC article.
References
-
- Sayed D., Abdellatif M.. Micrornas in development and disease. Physiol. Rev. 2011; 91:827–887. - PubMed
-
- Martin R.C., Liu P.P., Goloviznina N.A., Nonogaki H.. microRNA, seeds, and Darwin?: diverse function of miRNA in seed biology and plant responses to stress. J. Exp. Bot. 2010; 61:2229–2234. - PubMed
-
- Baulcombe D.C., Molnar A.. Crystal structure of p19–a universal suppressor of RNA silencing. Trends Biochem. Sci. 2004; 29:279–281. - PubMed
-
- Matzke M.A., Mosher R.A.. RNA-directed DNA methylation: an epigenetic pathway of increasing complexity. Nat. Rev. Genet. 2014; 15:394. - PubMed
-
- Bernstein E., Caudy A.A., Hammond S.M., Hannon G.J.. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001; 409:363–366. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
