

Enhancing auxin accumulation in maize root tips improves root growth and dwarfs plant height
- PMID: 28499064
- PMCID: PMC5785362
- DOI: 10.1111/pbi.12751
Enhancing auxin accumulation in maize root tips improves root growth and dwarfs plant height
Abstract
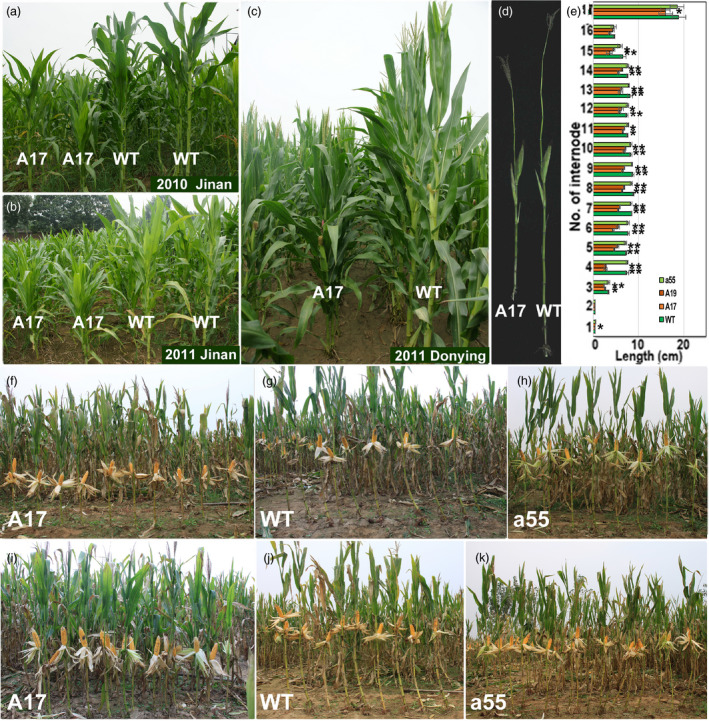
Maize is a globally important food, feed crop and raw material for the food and energy industry. Plant architecture optimization plays important roles in maize yield improvement. PIN-FORMED (PIN) proteins are important for regulating auxin spatiotemporal asymmetric distribution in multiple plant developmental processes. In this study, ZmPIN1a overexpression in maize increased the number of lateral roots and inhibited their elongation, forming a developed root system with longer seminal roots and denser lateral roots. ZmPIN1a overexpression reduced plant height, internode length and ear height. This modification of the maize phenotype increased the yield under high-density cultivation conditions, and the developed root system improved plant resistance to drought, lodging and a low-phosphate environment. IAA concentration, transport capacity determination and application of external IAA indicated that ZmPIN1a overexpression led to increased IAA transport from shoot to root. The increase in auxin in the root enabled the plant to allocate more carbohydrates to the roots, enhanced the growth of the root and improved plant resistance to environmental stress. These findings demonstrate that maize plant architecture can be improved by root breeding to create an ideal phenotype for further yield increases.
Keywords: ideal plant type; maize; root architecture modification; transgenic breeding; yield.
© 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures






Similar articles
-
Expression analysis of PIN-formed auxin efflux transporter genes in maize.Plant Signal Behav. 2019;14(9):1632689. doi: 10.1080/15592324.2019.1632689. Epub 2019 Jun 17. Plant Signal Behav. 2019. PMID: 31208285 Free PMC article.
-
Phosphate starvation of maize inhibits lateral root formation and alters gene expression in the lateral root primordium zone.BMC Plant Biol. 2012 Jun 14;12:89. doi: 10.1186/1471-2229-12-89. BMC Plant Biol. 2012. PMID: 22704465 Free PMC article.
-
Auxin efflux carrier ZmPIN1a modulates auxin reallocation involved in nitrate-mediated root formation.BMC Plant Biol. 2023 Feb 3;23(1):74. doi: 10.1186/s12870-023-04087-0. BMC Plant Biol. 2023. PMID: 36737696 Free PMC article.
-
Roles of auxin pathways in maize biology.J Exp Bot. 2023 Dec 1;74(22):6989-6999. doi: 10.1093/jxb/erad297. J Exp Bot. 2023. PMID: 37493143 Free PMC article. Review.
-
Genetic Control of Root System Development in Maize.Trends Plant Sci. 2018 Jan;23(1):79-88. doi: 10.1016/j.tplants.2017.10.004. Epub 2017 Nov 20. Trends Plant Sci. 2018. PMID: 29170008 Review.
Cited by
-
To Fight or to Grow: The Balancing Role of Ethylene in Plant Abiotic Stress Responses.Plants (Basel). 2021 Dec 23;11(1):33. doi: 10.3390/plants11010033. Plants (Basel). 2021. PMID: 35009037 Free PMC article. Review.
-
Comparative metabolomics of root-tips reveals distinct metabolic pathways conferring drought tolerance in contrasting genotypes of rice.BMC Genomics. 2023 Mar 27;24(1):152. doi: 10.1186/s12864-023-09246-z. BMC Genomics. 2023. PMID: 36973662 Free PMC article.
-
Modifications of Phytohormone Metabolism Aimed at Stimulation of Plant Growth, Improving Their Productivity and Tolerance to Abiotic and Biotic Stress Factors.Plants (Basel). 2022 Dec 8;11(24):3430. doi: 10.3390/plants11243430. Plants (Basel). 2022. PMID: 36559545 Free PMC article. Review.
-
Joint-GWAS, Linkage Mapping, and Transcriptome Analysis to Reveal the Genetic Basis of Plant Architecture-Related Traits in Maize.Int J Mol Sci. 2024 Feb 26;25(5):2694. doi: 10.3390/ijms25052694. Int J Mol Sci. 2024. PMID: 38473942 Free PMC article.
-
Genetic Basis of Seedling Root Traits in Common Wheat (Triticum aestivum L.) Identified by Genome-Wide Linkage Mapping.Plants (Basel). 2025 Feb 6;14(3):490. doi: 10.3390/plants14030490. Plants (Basel). 2025. PMID: 39943052 Free PMC article.
References
-
- Araus, J.L. , Slafer, G.A. , Royo, C. and Serret, M.D. (2008) Breeding for yield potential and stress adaptation in cereals. Crit. Rev. Plant Sci. 27, 377–412.
-
- Benkova, E. , Michniewicz, M. , Sauer, M. , Teichmann, T. , Seifertova, D. , Jurgens, G. and Friml, J. (2003) Local, efflux‐dependent auxin gradients as a common module for plant organ formation. Cell, 115, 591–602. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
