

MutMapPlus identified novel mutant alleles of a rice starch branching enzyme IIb gene for fine-tuning of cooked rice texture
- PMID: 28499068
- PMCID: PMC5785365
- DOI: 10.1111/pbi.12753
MutMapPlus identified novel mutant alleles of a rice starch branching enzyme IIb gene for fine-tuning of cooked rice texture
Abstract
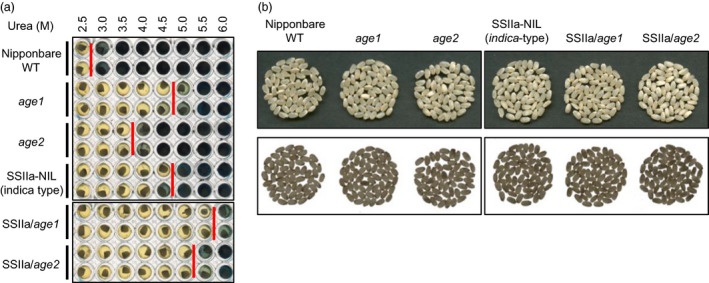
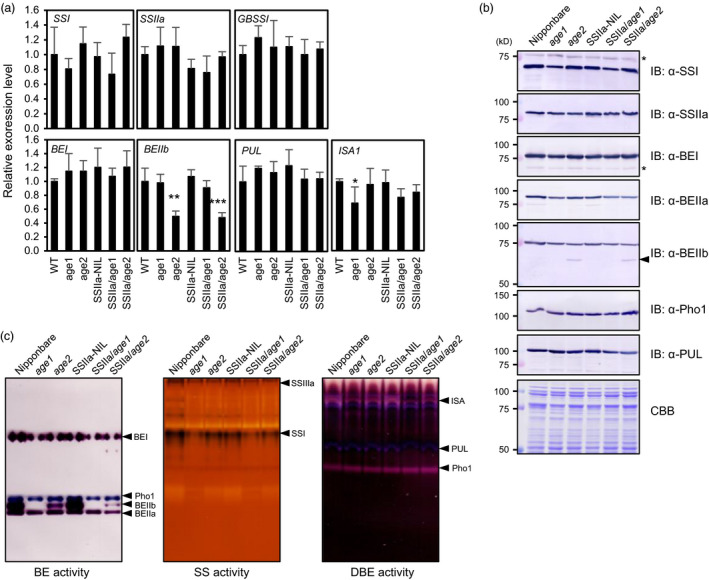
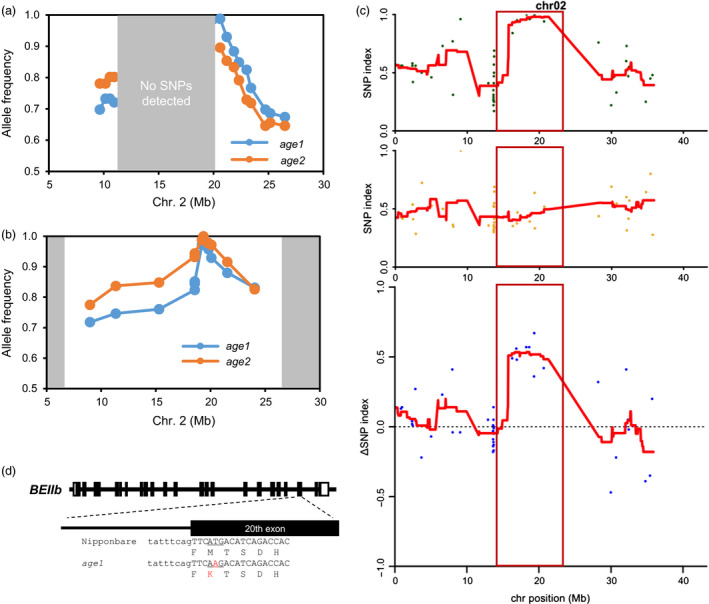
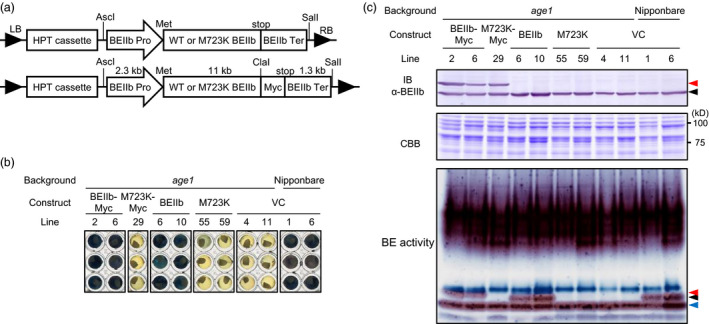
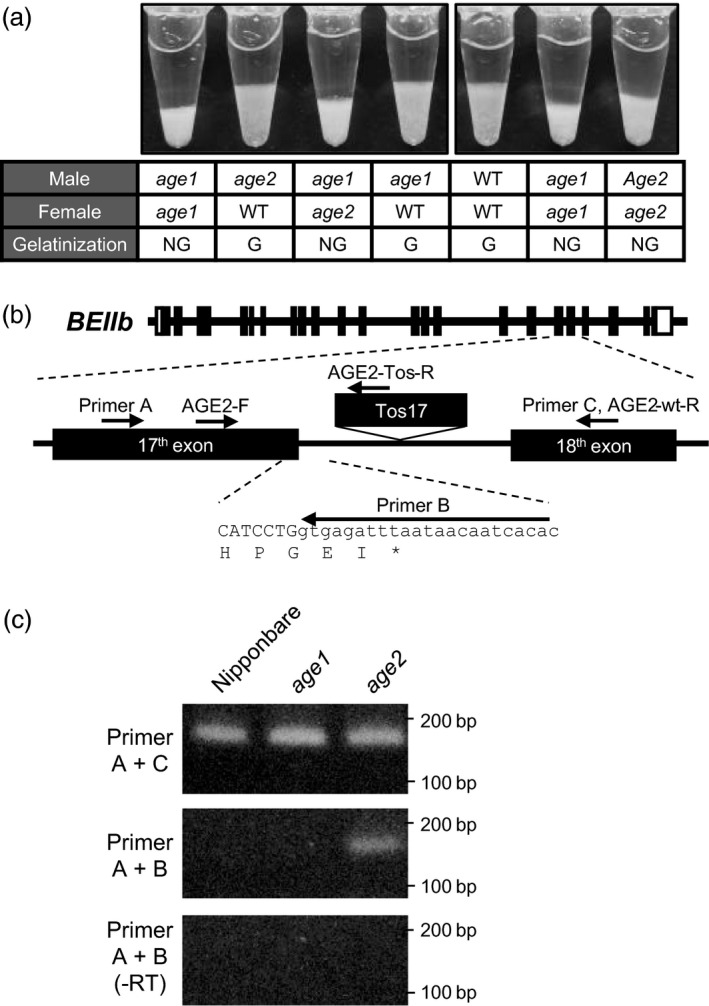
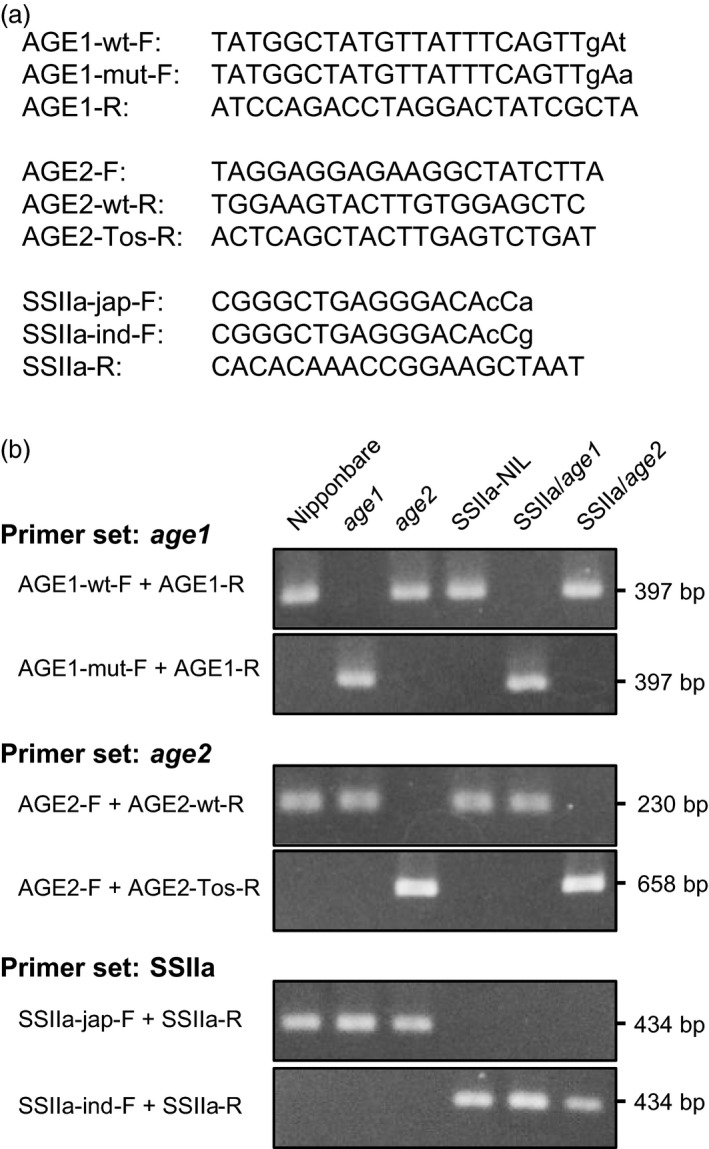
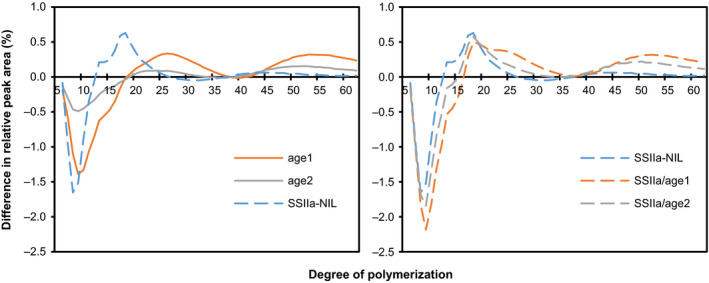
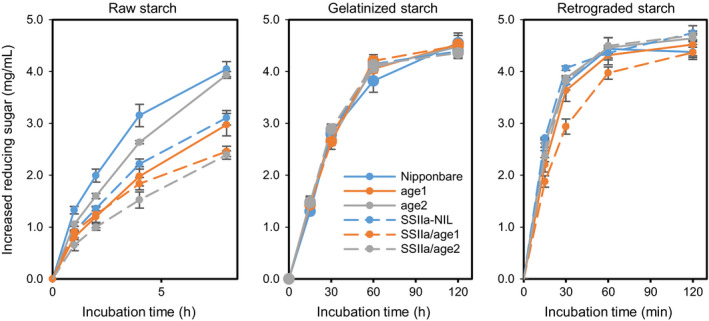
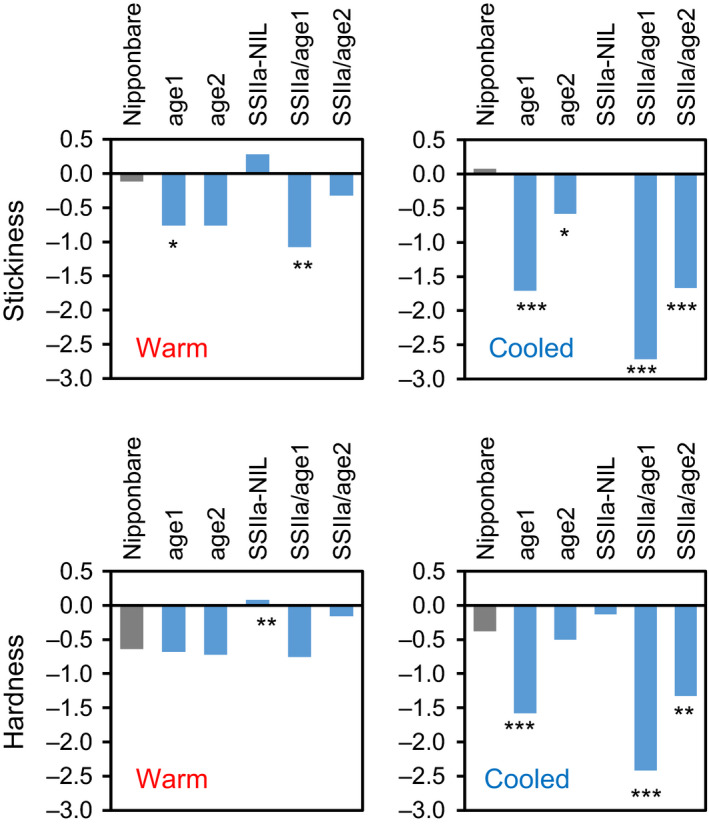
Physicochemical properties of storage starch largely determine rice grain quality and food characteristics. Therefore, modification of starch property is effective to fine-tune cooked rice textures. To obtain new resources with modified starch property as breeding materials, we screened a mutant population of a japonica cultivar Nipponbare and found two independent mutant lines, altered gelatinization (age)1 and age2, with moderate changes in starch gelatinization property. A combination of conventional genetic analyses and the latest mapping method, MutMapPlus, revealed that both of these lines harbour novel independent mutant alleles of starch branching enzyme IIb (BEIIb) gene. In age1, amino acid substitution of Met-723 to Lys completely abolished BEIIb enzyme activity without significant reduction in its protein level. A transposon insertion in an intron of BEIIb gene reduced BEIIb protein level and activity in age2. Production of a series of the mutant lines by combining age alleles and indica-type starch synthase IIa allele established stepwise alteration of the physicochemical properties of starch including apparent amylose content, thermal property, digestibility by α-amylase and branched structures of amylopectin. Consistent with the alteration of starch properties, the results of a sensory evaluation test demonstrated that warm cooked rice of the mutants showed a variety of textures without marked reduction in overall palatability. These results suggest that a series of the mutant lines are capable of manipulation of cooked rice textures.
Keywords: Oryza sativa; MutMapPlus; amylopectin; cooked rice texture; mutant allele; starch branching enzyme.
© 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures
Similar articles
-
Mutation in BEIIb mitigates the negative effect of the mutation in ISA1 on grain filling and amyloplast formation in rice.Plant Mol Biol. 2022 Mar;108(4-5):497-512. doi: 10.1007/s11103-022-01242-3. Epub 2022 Jan 27. Plant Mol Biol. 2022. PMID: 35083581
-
Relative importance of branching enzyme isoforms in determining starch fine structure and physicochemical properties of indica rice.Plant Mol Biol. 2022 Mar;108(4-5):399-412. doi: 10.1007/s11103-021-01207-y. Epub 2021 Nov 8. Plant Mol Biol. 2022. PMID: 34750721
-
The role of different Wx and BEIIb allele combinations on fine structures and functional properties of indica rice starches.Carbohydr Polym. 2022 Feb 15;278:118972. doi: 10.1016/j.carbpol.2021.118972. Epub 2021 Dec 5. Carbohydr Polym. 2022. PMID: 34973786
-
Starch fine molecular structures: The basis for designer rice with slower digestibility and desirable texture properties.Carbohydr Polym. 2023 Jan 1;299:120217. doi: 10.1016/j.carbpol.2022.120217. Epub 2022 Oct 13. Carbohydr Polym. 2023. PMID: 36876819 Review.
-
Towards a better understanding of the metabolic system for amylopectin biosynthesis in plants: rice endosperm as a model tissue.Plant Cell Physiol. 2002 Jul;43(7):718-25. doi: 10.1093/pcp/pcf091. Plant Cell Physiol. 2002. PMID: 12154134 Review.
Cited by
-
SUPER STARCHY1/ONAC025 participates in rice grain filling.Plant Direct. 2020 Sep 4;4(9):e00249. doi: 10.1002/pld3.249. eCollection 2020 Sep. Plant Direct. 2020. PMID: 32995698 Free PMC article.
-
Discrepancies in resistant starch and starch physicochemical properties between rice mutants similar in high amylose content.Front Plant Sci. 2023 Nov 3;14:1267281. doi: 10.3389/fpls.2023.1267281. eCollection 2023. Front Plant Sci. 2023. PMID: 38023836 Free PMC article.
-
Regulation of Amylose Content by Single Mutations at an Active Site in the Wx-B1 Gene in a Tetraploid Wheat Mutant.Int J Mol Sci. 2022 Jul 29;23(15):8432. doi: 10.3390/ijms23158432. Int J Mol Sci. 2022. PMID: 35955567 Free PMC article.
-
Genetic region responsible for the differences of starch properties in two glutinous rice cultivars in Hokkaido, Japan.Breed Sci. 2021 Jun;71(3):375-383. doi: 10.1270/jsbbs.20163. Epub 2021 Jun 25. Breed Sci. 2021. PMID: 34776744 Free PMC article.
-
Next Generation Sequencing Based Forward Genetic Approaches for Identification and Mapping of Causal Mutations in Crop Plants: A Comprehensive Review.Plants (Basel). 2020 Oct 14;9(10):1355. doi: 10.3390/plants9101355. Plants (Basel). 2020. PMID: 33066352 Free PMC article. Review.
References
-
- Abe, A. , Kosugi, S. , Yoshida, K. , Natsume, S. , Takagi, H. , Kanzaki, H. , Matsumura, H. et al. (2012) Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 30, 174–178. - PubMed
-
- Ahmed, Z. , Tetlow, I.J. , Ahmed, R. , Morell, M.K. and Emes, M.J. (2015) Protein‐protein interactions among enzymes of starch biosynthesis in high‐amylose barley genotypes reveal differential roles of heteromeric enzyme complexes in the synthesis of A and B granules. Plant Sci. 233, 95–106. - PubMed
-
- Ball, S.G. and Morell, M.K. (2003) From bacterial glycogen to starch: understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol. 54, 207–233. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
