

Acyl-CoA-Binding Protein ACBP1 Modulates Sterol Synthesis during Embryogenesis
- PMID: 28500265
- PMCID: PMC5490911
- DOI: 10.1104/pp.17.00412
Acyl-CoA-Binding Protein ACBP1 Modulates Sterol Synthesis during Embryogenesis
Abstract
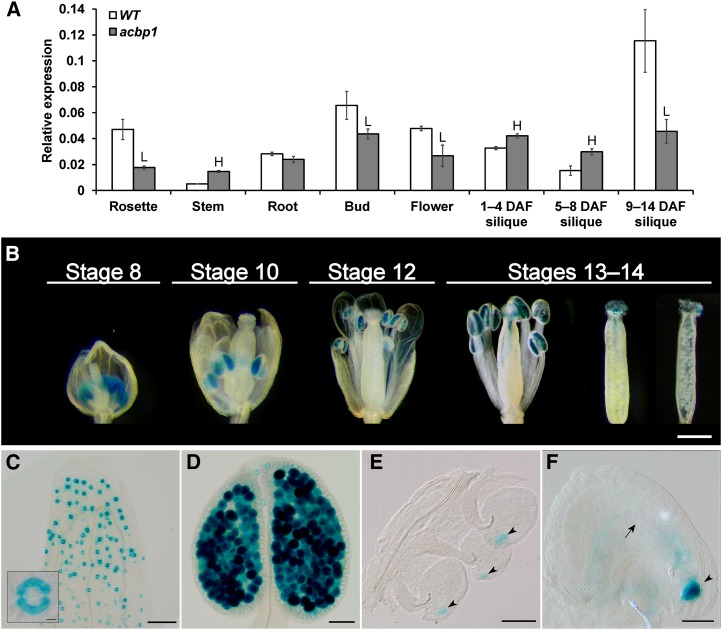
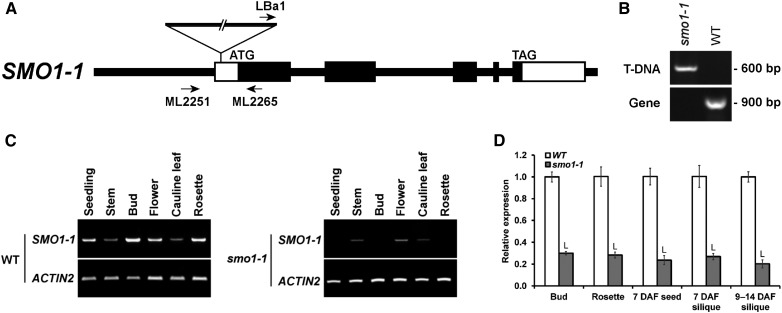
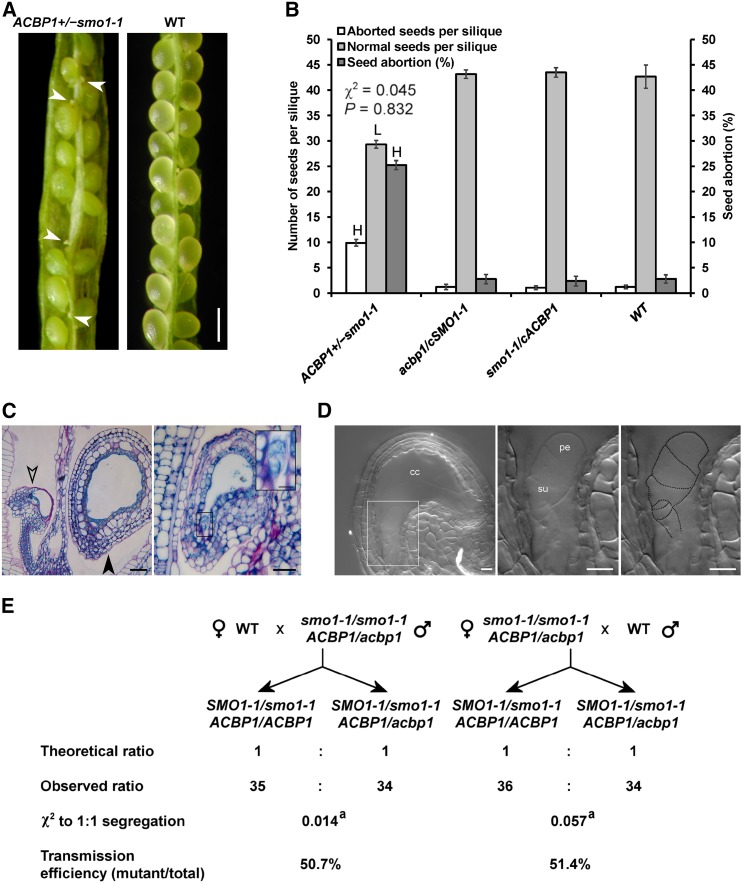
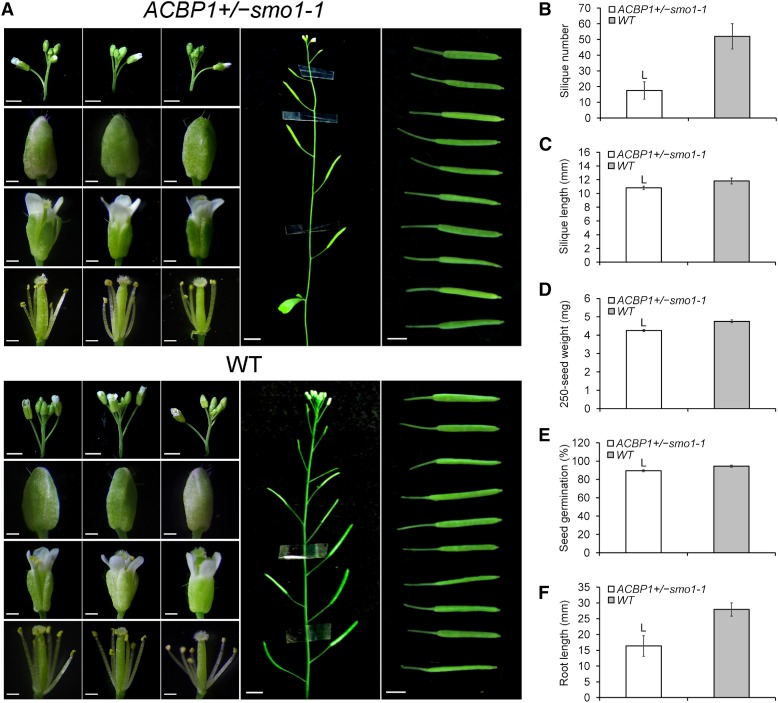
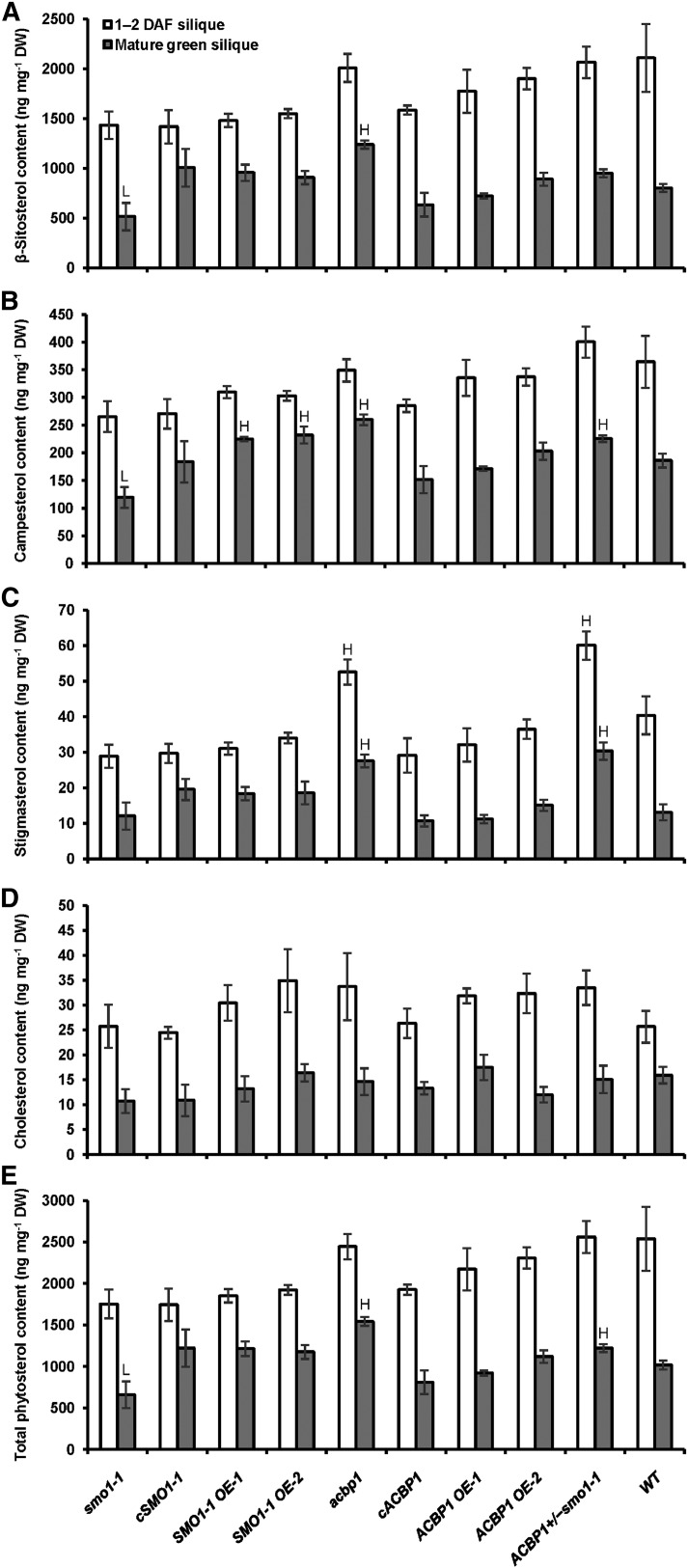
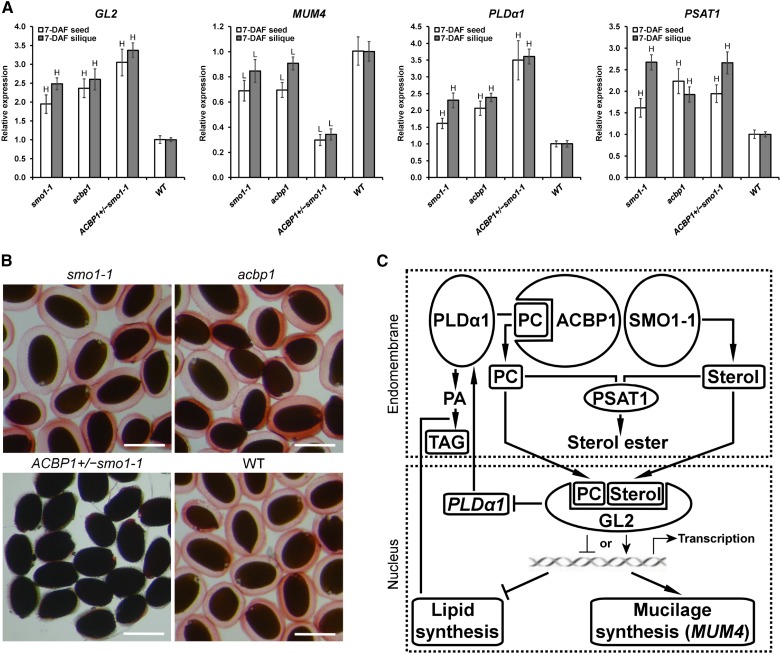
Fatty acids (FAs) and sterols are primary metabolites that exert interrelated functions as structural and signaling lipids. Despite their common syntheses from acetyl-coenzyme A, homeostatic cross talk remains enigmatic. Six Arabidopsis (Arabidopsis thaliana) acyl-coenzyme A-binding proteins (ACBPs) are involved in FA metabolism. ACBP1 interacts with PHOSPHOLIPASE Dα1 and regulates phospholipid composition. Here, its specific role in the negative modulation of sterol synthesis during embryogenesis is reported. ACBP1, likely in a liganded state, interacts with STEROL C4-METHYL OXIDASE1-1 (SMO1-1), a rate-limiting enzyme in the sterol pathway. Proembryo abortion in the double mutant indicated that the ACBP1-SMO1-1 interaction is synthetic lethal, corroborating with their strong promoter activities in developing ovules. Gas chromatography-mass spectrometry revealed quantitative and compositional changes in FAs and sterols upon overexpression or mutation of ACBP1 and/or SMO1-1 Aberrant levels of these metabolites may account for the downstream defect in lipid signaling. GLABRA2 (GL2), encoding a phospholipid/sterol-binding homeodomain transcription factor, was up-regulated in developing seeds of acbp1, smo1-1, and ACBP1+/-smo1-1 in comparison with the wild type. Consistent with the corresponding transcriptional alteration of GL2 targets, high-oil, low-mucilage phenotypes of gl2 were phenocopied in ACBP1+/-smo1-1 Thus, ACBP1 appears to modulate the metabolism of two important lipid classes (FAs and sterols) influencing cellular signaling.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures

Similar articles
-
Arabidopsis ACYL-COA-BINDING PROTEIN1 interacts with STEROL C4-METHYL OXIDASE1-2 to modulate gene expression of homeodomain-leucine zipper IV transcription factors.New Phytol. 2018 Apr;218(1):183-200. doi: 10.1111/nph.14965. Epub 2017 Dec 30. New Phytol. 2018. PMID: 29288621
-
The Arabidopsis acbp1acbp2 double mutant lacking acyl-CoA-binding proteins ACBP1 and ACBP2 is embryo lethal.New Phytol. 2010 Jun;186(4):843-855. doi: 10.1111/j.1469-8137.2010.03231.x. Epub 2010 Mar 22. New Phytol. 2010. PMID: 20345632 Free PMC article.
-
Overexpression of membrane-associated acyl-CoA-binding protein ACBP1 enhances lead tolerance in Arabidopsis.Plant J. 2008 Apr;54(1):141-51. doi: 10.1111/j.1365-313X.2008.03402.x. Epub 2008 Jan 7. Plant J. 2008. PMID: 18182029
-
An Arabidopsis family of six acyl-CoA-binding proteins has three cytosolic members.Plant Physiol Biochem. 2009 Jun;47(6):479-84. doi: 10.1016/j.plaphy.2008.12.002. Epub 2008 Dec 16. Plant Physiol Biochem. 2009. PMID: 19121948 Review.
-
Deciphering the roles of acyl-CoA-binding proteins in plant cells.Protoplasma. 2016 Sep;253(5):1177-95. doi: 10.1007/s00709-015-0882-6. Epub 2015 Sep 4. Protoplasma. 2016. PMID: 26340904 Review.
Cited by
-
Overexpression of HMG-CoA synthase promotes Arabidopsis root growth and adversely affects glucosinolate biosynthesis.J Exp Bot. 2020 Jan 1;71(1):272-289. doi: 10.1093/jxb/erz420. J Exp Bot. 2020. PMID: 31557302 Free PMC article.
-
Stigmasterol: An Enigmatic Plant Stress Sterol with Versatile Functions.Int J Mol Sci. 2024 Jul 25;25(15):8122. doi: 10.3390/ijms25158122. Int J Mol Sci. 2024. PMID: 39125690 Free PMC article. Review.
-
Depletion of Arabidopsis ACYL-COA-BINDING PROTEIN3 Affects Fatty Acid Composition in the Phloem.Front Plant Sci. 2018 Jan 25;9:2. doi: 10.3389/fpls.2018.00002. eCollection 2018. Front Plant Sci. 2018. PMID: 29422909 Free PMC article.
-
Interaction of Soybean (Glycine max (L.) Merr.) Class II ACBPs with MPK2 and SAPK2 Kinases: New Insights into the Regulatory Mechanisms of Plant ACBPs.Plants (Basel). 2024 Apr 19;13(8):1146. doi: 10.3390/plants13081146. Plants (Basel). 2024. PMID: 38674555 Free PMC article.
-
Metabolism and Biological Activities of 4-Methyl-Sterols.Molecules. 2019 Jan 27;24(3):451. doi: 10.3390/molecules24030451. Molecules. 2019. PMID: 30691248 Free PMC article. Review.
References
-
- Bach TJ. (1995) Some new aspects of isoprenoid biosynthesis in plants: a review. Lipids 30: 191–202 - PubMed
-
- Beh CT, Rine J (2004) A role for yeast oxysterol-binding protein homologs in endocytosis and in the maintenance of intracellular sterol-lipid distribution. J Cell Sci 117: 2983–2996 - PubMed
-
- Benveniste P. (2004) Biosynthesis and accumulation of sterols. Annu Rev Plant Biol 55: 429–457 - PubMed
-
- Block MA, Dorne AJ, Joyard J, Douce R (1983) The acyl-CoA synthetase and acyl-CoA thioesterase are located on the outer and inner membrane of the chloroplast envelope, respectively. FEBS Lett 153: 377–381
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
