

Cytoskeletal Proteins in Caulobacter crescentus: Spatial Orchestrators of Cell Cycle Progression, Development, and Cell Shape
- PMID: 28500524
- PMCID: PMC5554946
- DOI: 10.1007/978-3-319-53047-5_4
Cytoskeletal Proteins in Caulobacter crescentus: Spatial Orchestrators of Cell Cycle Progression, Development, and Cell Shape
Abstract
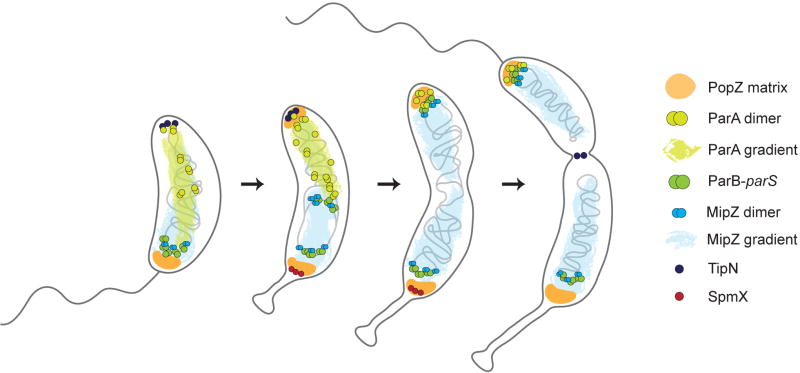
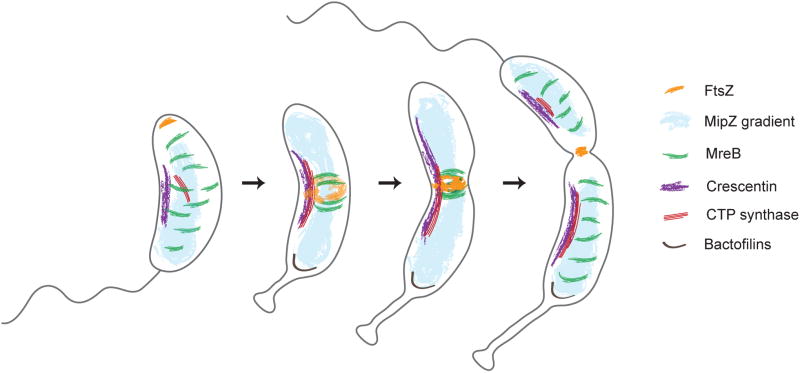
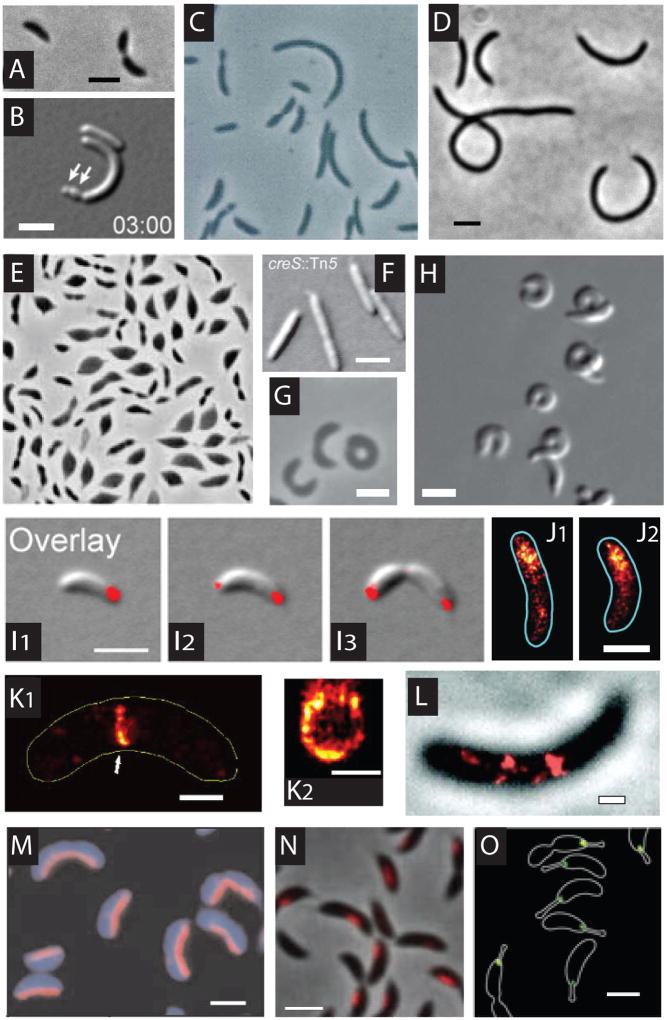
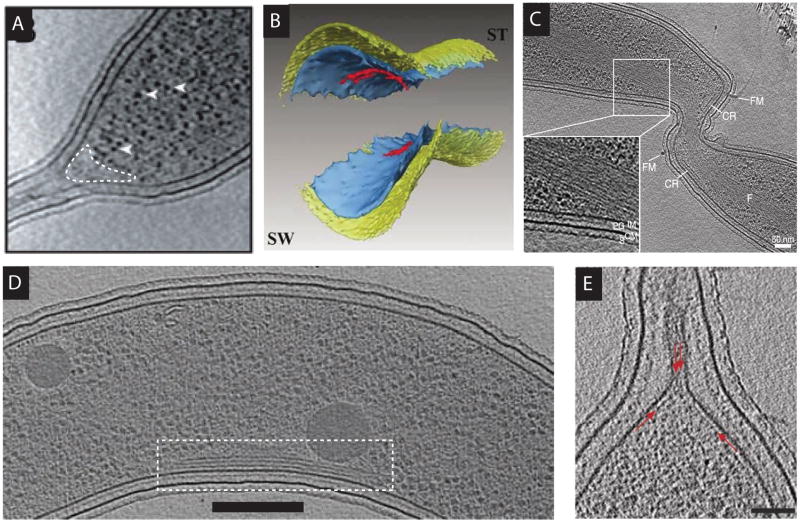
Caulobacter crescentus, an aquatic Gram-negative α-proteobacterium, is dimorphic, as a result of asymmetric cell divisions that give rise to a free-swimming swarmer daughter cell and a stationary stalked daughter. Cell polarity of vibrioid C. crescentus cells is marked by the presence of a stalk at one end in the stationary form and a polar flagellum in the motile form. Progression through the cell cycle and execution of the associated morphogenetic events are tightly controlled through regulation of the abundance and activity of key proteins. In synergy with the regulation of protein abundance or activity, cytoskeletal elements are key contributors to cell cycle progression through spatial regulation of developmental processes. These include: polarity establishment and maintenance, DNA segregation, cytokinesis, and cell elongation. Cytoskeletal proteins in C. crescentus are additionally required to maintain its rod shape, curvature, and pole morphology. In this chapter, we explore the mechanisms through which cytoskeletal proteins in C. crescentus orchestrate developmental processes by acting as scaffolds for protein recruitment, generating force, and/or restricting or directing the motion of molecular machines. We discuss each cytoskeletal element in turn, beginning with those important for organization of molecules at the cell poles and chromosome segregation, then cytokinesis, and finally cell shape.
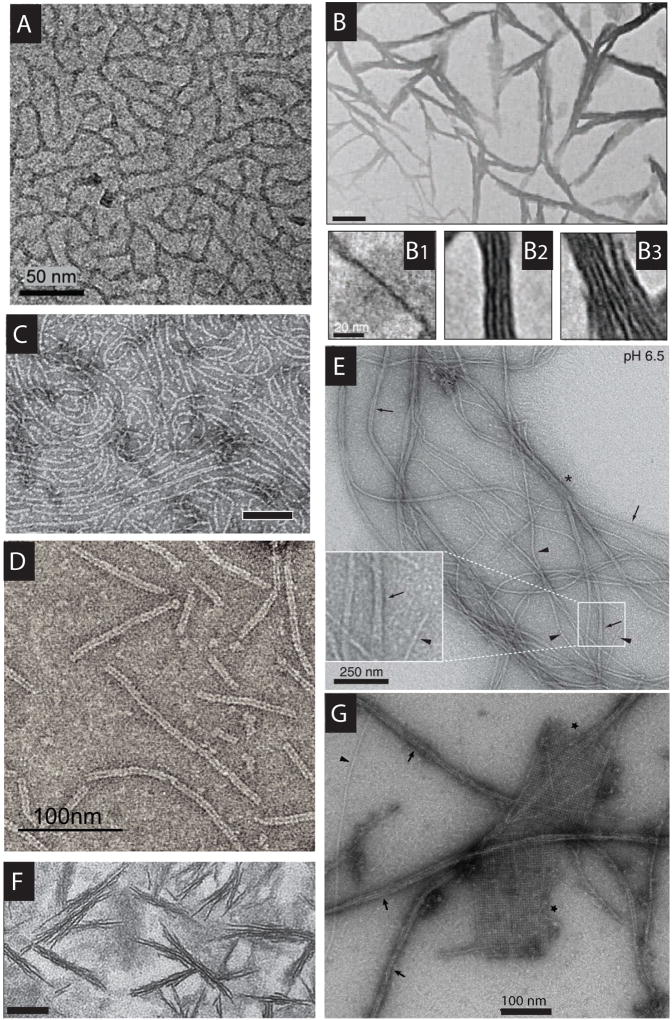
Figures
Similar articles
-
Shapeshifting to Survive: Shape Determination and Regulation in Caulobacter crescentus.Trends Microbiol. 2017 Aug;25(8):673-687. doi: 10.1016/j.tim.2017.03.006. Epub 2017 Mar 27. Trends Microbiol. 2017. PMID: 28359631 Free PMC article. Review.
-
Cell cycle regulation in Caulobacter: location, location, location.J Cell Sci. 2007 Oct 15;120(Pt 20):3501-7. doi: 10.1242/jcs.005967. J Cell Sci. 2007. PMID: 17928306 Review.
-
Cell cycle and positional constraints on FtsZ localization and the initiation of cell division in Caulobacter crescentus.Mol Microbiol. 2001 Feb;39(4):949-59. doi: 10.1046/j.1365-2958.2001.02287.x. Mol Microbiol. 2001. PMID: 11251815
-
The Caulobacter crescentus polar organelle development protein PodJ is differentially localized and is required for polar targeting of the PleC development regulator.Mol Microbiol. 2003 Feb;47(4):929-41. doi: 10.1046/j.1365-2958.2003.03349.x. Mol Microbiol. 2003. PMID: 12581350
-
The chromosome partitioning protein, ParB, is required for cytokinesis in Caulobacter crescentus.Mol Microbiol. 2001 Nov;42(3):741-55. doi: 10.1046/j.1365-2958.2001.02643.x. Mol Microbiol. 2001. PMID: 11722739
Cited by
-
Structural Determinants and Their Role in Cyanobacterial Morphogenesis.Life (Basel). 2020 Dec 17;10(12):355. doi: 10.3390/life10120355. Life (Basel). 2020. PMID: 33348886 Free PMC article. Review.
-
A bacterial cytolinker couples positioning of magnetic organelles to cell shape control.Proc Natl Acad Sci U S A. 2020 Dec 15;117(50):32086-32097. doi: 10.1073/pnas.2014659117. Epub 2020 Nov 30. Proc Natl Acad Sci U S A. 2020. PMID: 33257551 Free PMC article.
-
Epiplasts: Membrane Skeletons and Epiplastin Proteins in Euglenids, Glaucophytes, Cryptophytes, Ciliates, Dinoflagellates, and Apicomplexans.mBio. 2018 Oct 30;9(5):e02020-18. doi: 10.1128/mBio.02020-18. mBio. 2018. PMID: 30377285 Free PMC article.
-
FzlA, an essential regulator of FtsZ filament curvature, controls constriction rate during Caulobacter division.Mol Microbiol. 2018 Jan;107(2):180-197. doi: 10.1111/mmi.13876. Epub 2017 Dec 1. Mol Microbiol. 2018. PMID: 29119622 Free PMC article.
-
FtsEX-mediated regulation of the final stages of cell division reveals morphogenetic plasticity in Caulobacter crescentus.PLoS Genet. 2017 Sep 8;13(9):e1006999. doi: 10.1371/journal.pgen.1006999. eCollection 2017 Sep. PLoS Genet. 2017. PMID: 28886022 Free PMC article.
References
-
- Aaron M, et al. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Molecular Microbiology. 2007;64(4):938–952. - PubMed
-
- Ausmees N, Kuhn JR, Jacobs-Wagner C. The bacterial cytoskeleton: an intermediate filament-like function in cell shape. Cell. 2003;115(6):705–713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
