

Butyrate reduces high-fat diet-induced metabolic alterations, hepatic steatosis and pancreatic beta cell and intestinal barrier dysfunctions in prediabetic mice
- PMID: 28504618
- PMCID: PMC5476343
- DOI: 10.1177/1535370217708188
Butyrate reduces high-fat diet-induced metabolic alterations, hepatic steatosis and pancreatic beta cell and intestinal barrier dysfunctions in prediabetic mice
Abstract
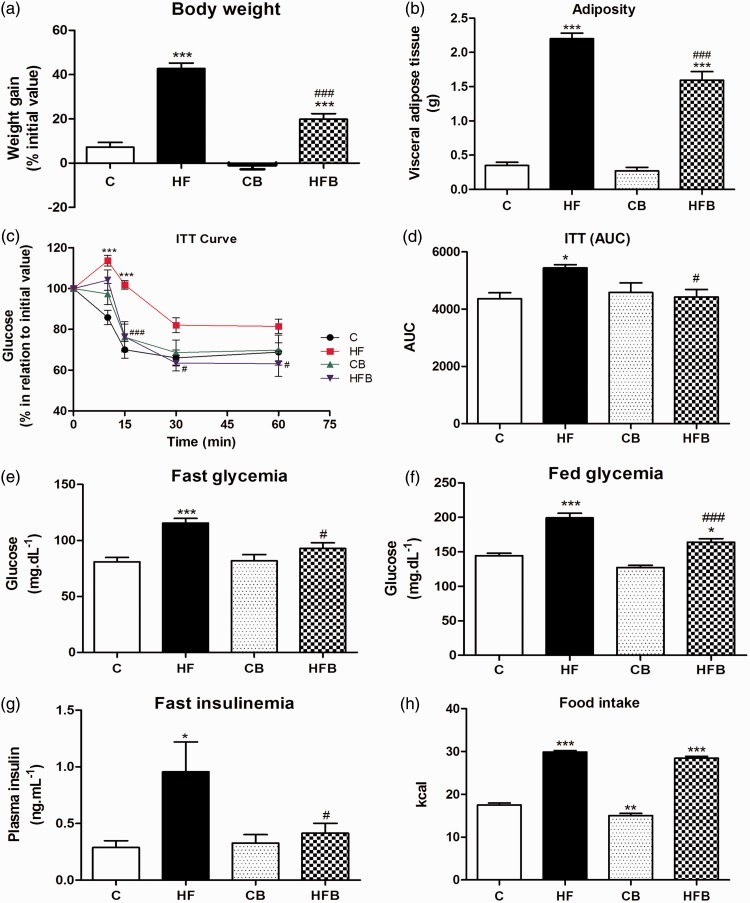
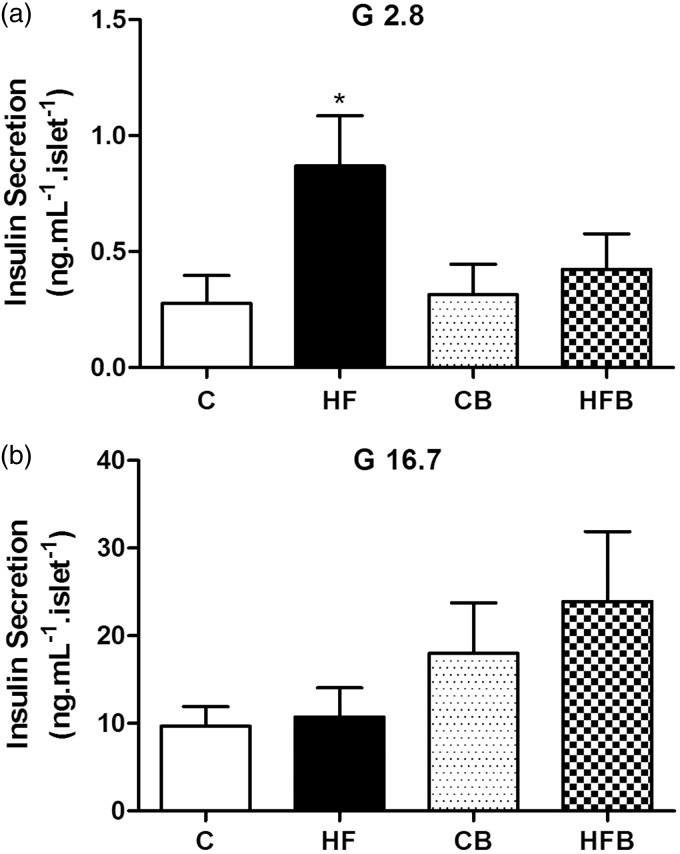
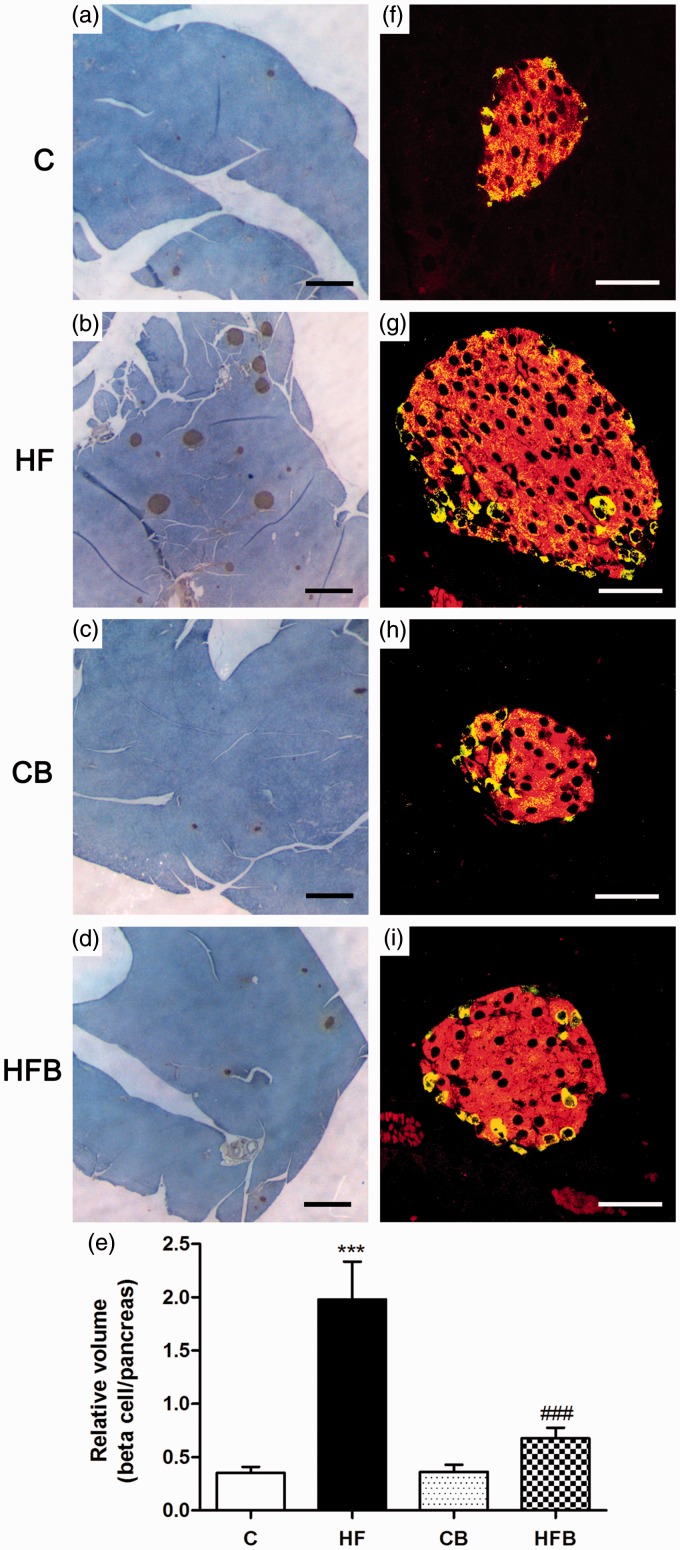
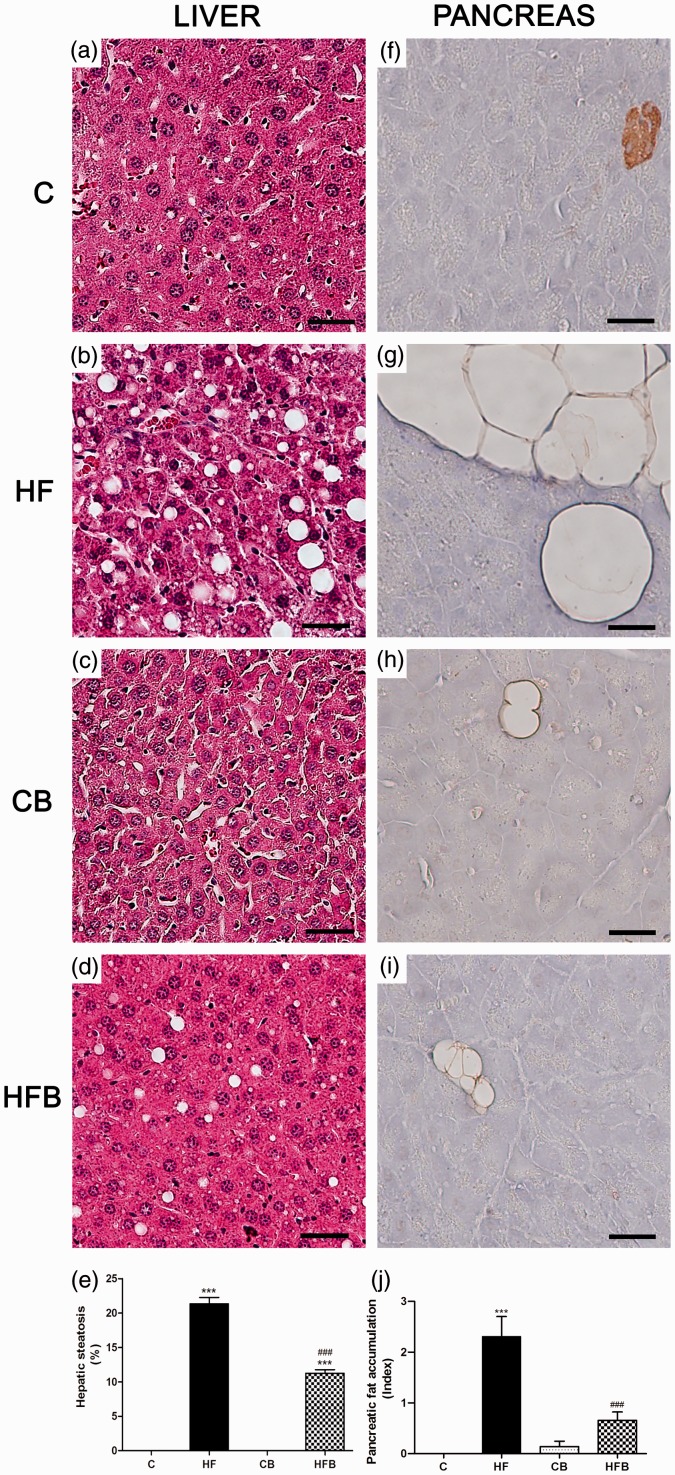
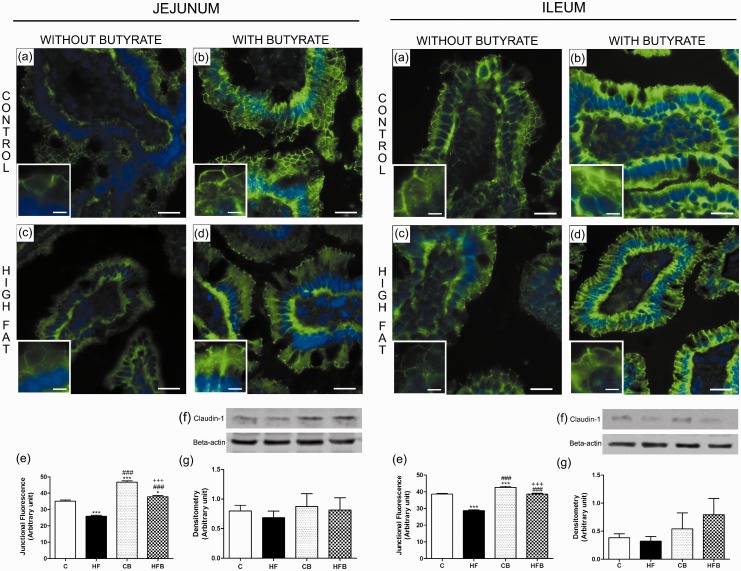
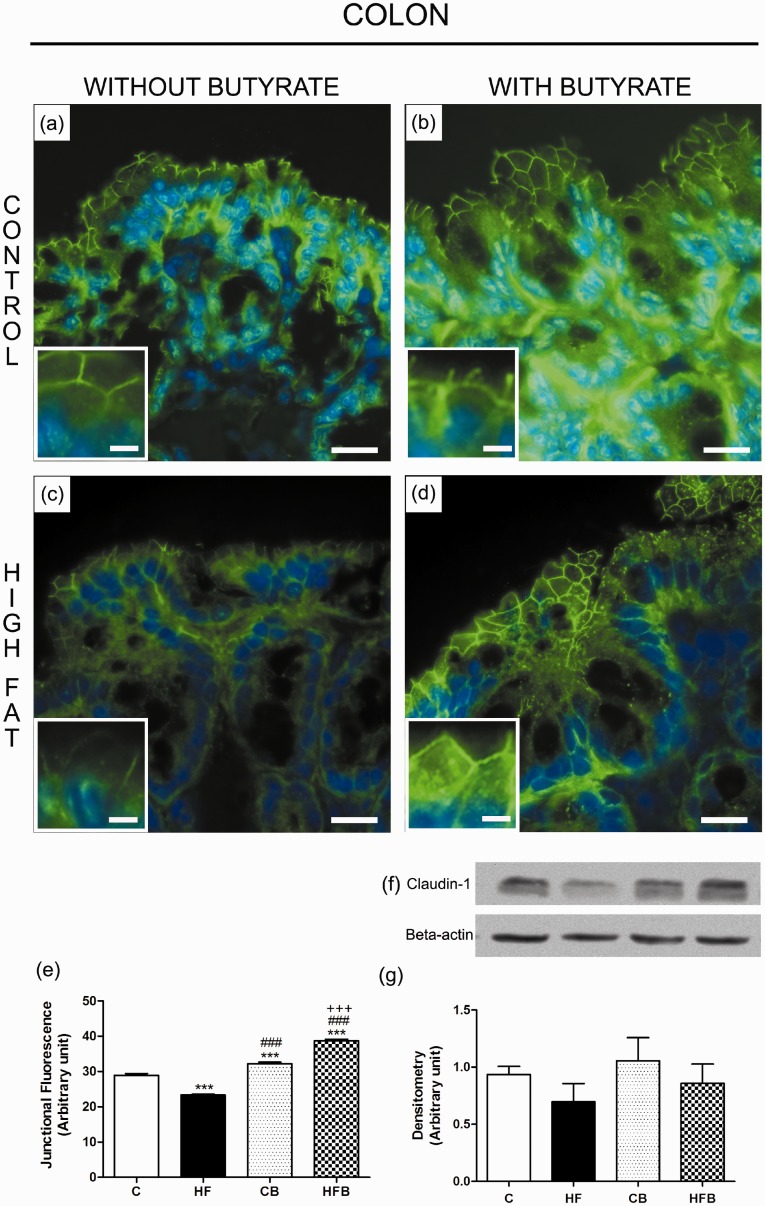
In this study, we investigated the effect of diet supplementation with sodium butyrate (5% w/w), a short-chain fatty acid produced by the intestinal microbiota, on metabolic parameters, body adiposity, hepatic and pancreatic lipid accumulation, beta cell function/mass as well as on the structure and function of the tight junction-mediated intestinal epithelial barrier in both normal and obese/prediabetic C57 mice fed a regular (control) or high-fat diet for 60 days, respectively. Butyrate treatment significantly inhibited all the high-fat-induced metabolic dysfunctions evaluated, i.e. significantly reduced the weight gain and body adiposity as well as the insulin resistant state, hyperglycemia and hyperinsulinemia, without changing food intake. In addition, high-fat-fed mice treated with this short-chain fatty acid displayed no compensatory hyperplasia of pancreatic beta cells nor marked hepatic steatosis as seen in prediabetic mice after high-fat diet only. Isolated pancreatic islets from high-fat-fed mice treated with butyrate showed improvement of the insulin secretion, which was associated with a significant decrease in lipid accumulation within the pancreas. Butyrate enhanced the intestinal epithelial barrier, as revealed by the FITC-Dextran permeability assay, which was accompanied by a significant increase in the junctional content of the tight junction-associated claudin-1 in intestinal epithelia of jejunum, ileum, and colon of both control and high-fat mice. In conclusion, our results showed that diet supplementation with butyrate inhibits the deleterious effects of high-fat diet intake on metabolic parameters and structure/function of several tissues/organs associated with type 2 diabetes mellitus in a mouse model, suggesting a potential use of this short-chain fatty acid in the treatment of this endocrine-metabolic disorder. Impact statement Butyrate is a short-chain fatty acid produced by the intestinal microbiota through the fermentation of non-absorbable carbohydrates and proteins (e.g. fibers). Sodium butyrate incorporated into the diet displayed a protective action on metabolic, hepatic, pancreatic and intestinal alterations induced by high-fat diet in mice, resulting in significant inhibition of the development of a prediabetic state. Thus, our data suggest that butyrate may have a potential therapeutic use in the treatment of type 2 diabetes and related disorders.
Keywords: Butyrate; high-fat diet; intestinal paracellular barrier; obesity; pancreatic beta cell; type 2 diabetes mellitus.
Figures
References
-
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diab Care 2014; 37: 81–90.
-
- Thomas CC, Philipson LH. Update on diabetes classification. Med Clin North Am 2015; 99: 1–16. - PubMed
-
- Canivell S, Gomis R. Diagnosis and classification of autoimmune diabetes mellitus. Autoimmun Rev 2014; 13: 403–7. - PubMed
-
- Tripathy D, Chavez AO. Defects in insulin secretion and action in the pathogenesis of type 2 diabetes mellitus. Curr Diab Rep 2010; 10: 184–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical