

PGBD5 promotes site-specific oncogenic mutations in human tumors
- PMID: 28504702
- PMCID: PMC5489359
- DOI: 10.1038/ng.3866
PGBD5 promotes site-specific oncogenic mutations in human tumors
Erratum in
-
Erratum: PGBD5 promotes site-specific oncogenic mutations in human tumors.Nat Genet. 2017 Sep 27;49(10):1558. doi: 10.1038/ng1017-1558b. Nat Genet. 2017. PMID: 28951624 No abstract available.
-
Author Correction: PGBD5 promotes site-specific oncogenic mutations in human tumors.Nat Genet. 2020 Nov;52(11):1265. doi: 10.1038/s41588-020-00711-z. Nat Genet. 2020. PMID: 32918070
Abstract
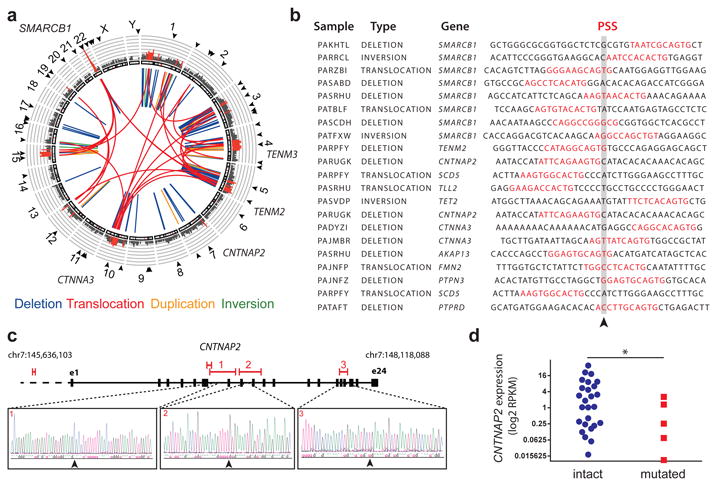
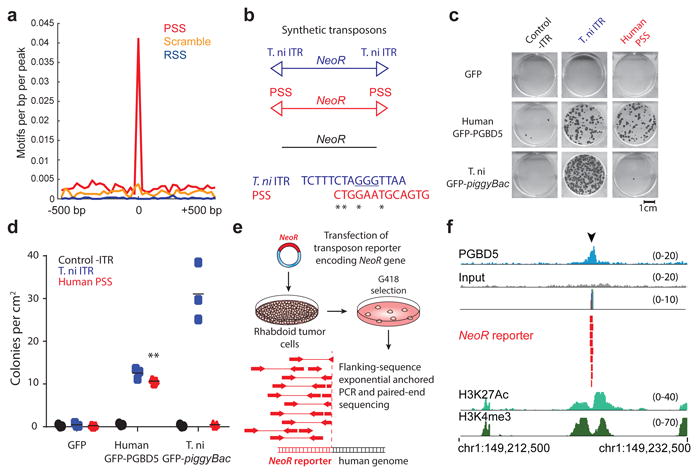
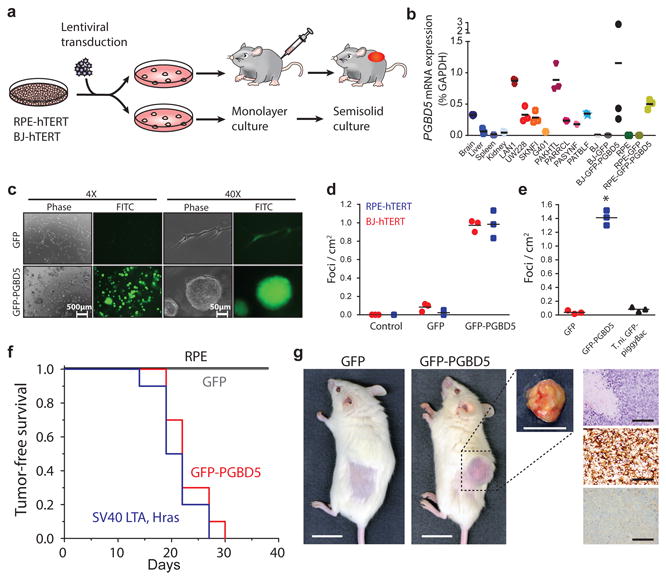
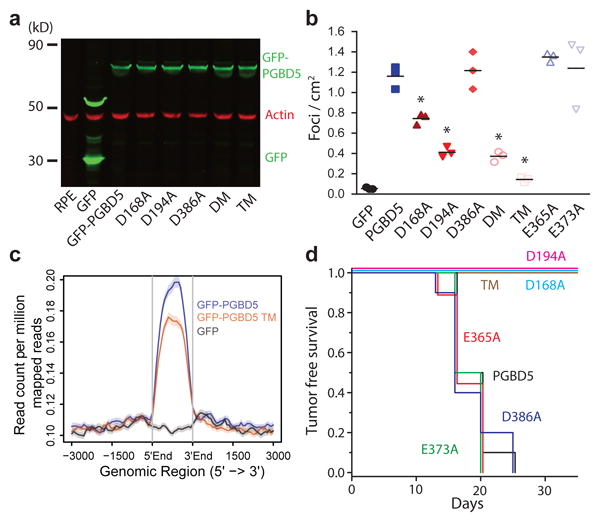
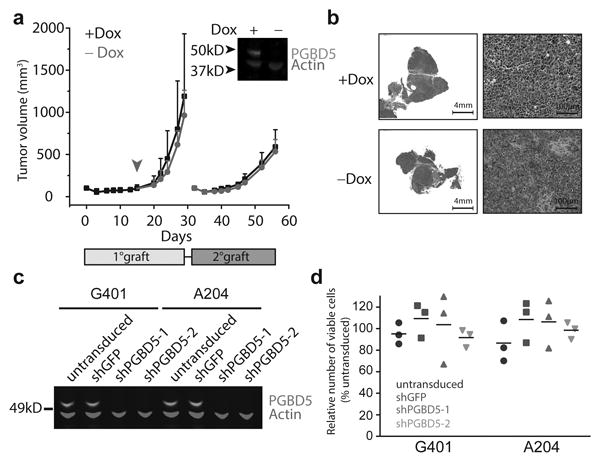
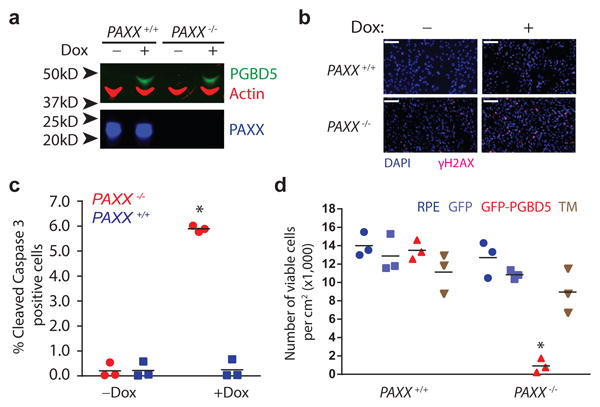
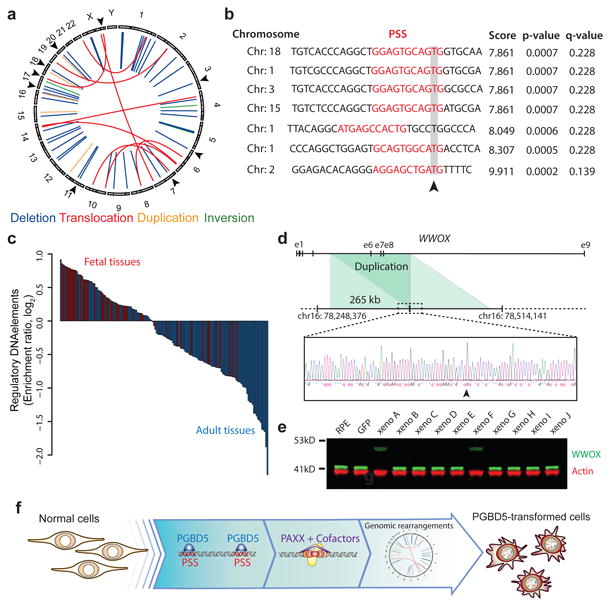
Genomic rearrangements are a hallmark of human cancers. Here, we identify the piggyBac transposable element derived 5 (PGBD5) gene as encoding an active DNA transposase expressed in the majority of childhood solid tumors, including lethal rhabdoid tumors. Using assembly-based whole-genome DNA sequencing, we found previously undefined genomic rearrangements in human rhabdoid tumors. These rearrangements involved PGBD5-specific signal (PSS) sequences at their breakpoints and recurrently inactivated tumor-suppressor genes. PGBD5 was physically associated with genomic PSS sequences that were also sufficient to mediate PGBD5-induced DNA rearrangements in rhabdoid tumor cells. Ectopic expression of PGBD5 in primary immortalized human cells was sufficient to promote cell transformation in vivo. This activity required specific catalytic residues in the PGBD5 transposase domain as well as end-joining DNA repair and induced structural rearrangements with PSS breakpoints. These results define PGBD5 as an oncogenic mutator and provide a plausible mechanism for site-specific DNA rearrangements in childhood and adult solid tumors.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
