

Wounding induces dedifferentiation of epidermal Gata6+ cells and acquisition of stem cell properties
- PMID: 28504705
- PMCID: PMC7610974
- DOI: 10.1038/ncb3532
Wounding induces dedifferentiation of epidermal Gata6+ cells and acquisition of stem cell properties
Abstract
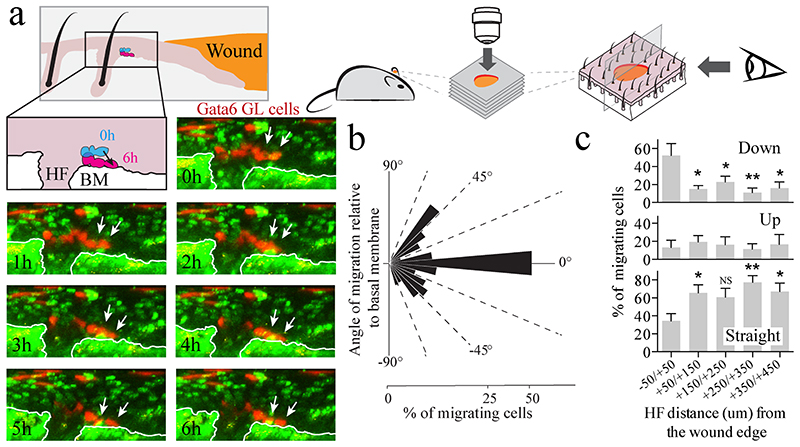
The epidermis is maintained by multiple stem cell populations whose progeny differentiate along diverse, and spatially distinct, lineages. Here we show that the transcription factor Gata6 controls the identity of the previously uncharacterized sebaceous duct (SD) lineage and identify the Gata6 downstream transcription factor network that specifies a lineage switch between sebocytes and SD cells. During wound healing differentiated Gata6+ cells migrate from the SD into the interfollicular epidermis and dedifferentiate, acquiring the ability to undergo long-term self-renewal and differentiate into a much wider range of epidermal lineages than in undamaged tissue. Our data not only demonstrate that the structural and functional complexity of the junctional zone is regulated by Gata6, but also reveal that dedifferentiation is a previously unrecognized property of post-mitotic, terminally differentiated cells that have lost contact with the basement membrane. This resolves the long-standing debate about the contribution of terminally differentiated cells to epidermal wound repair.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
