

Spatially restricted dental regeneration drives pufferfish beak development
- PMID: 28507130
- PMCID: PMC5465925
- DOI: 10.1073/pnas.1702909114
Spatially restricted dental regeneration drives pufferfish beak development
Abstract
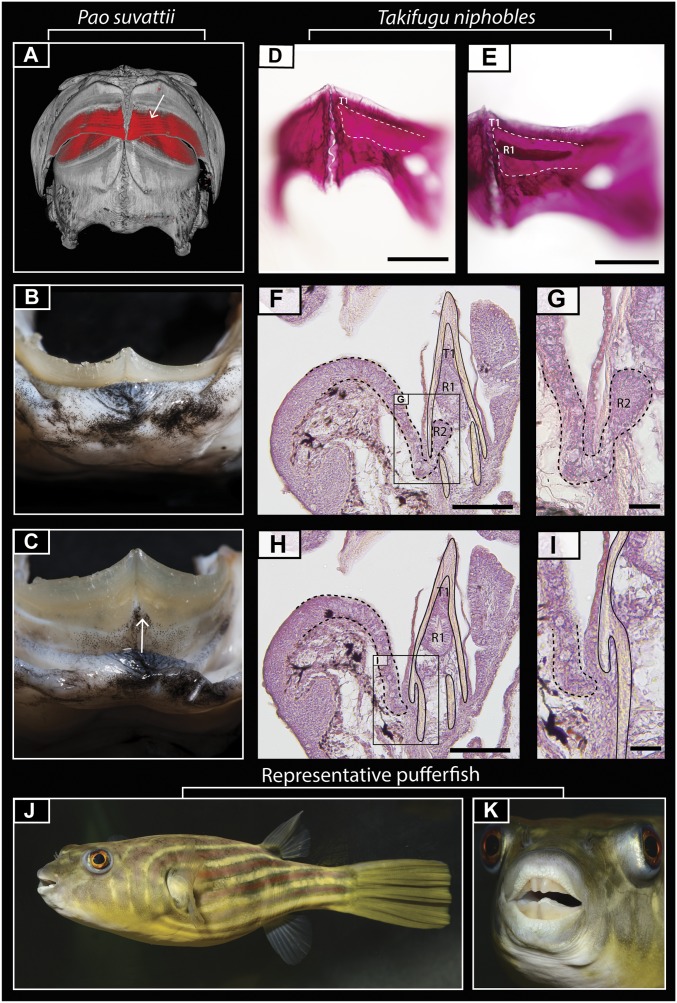
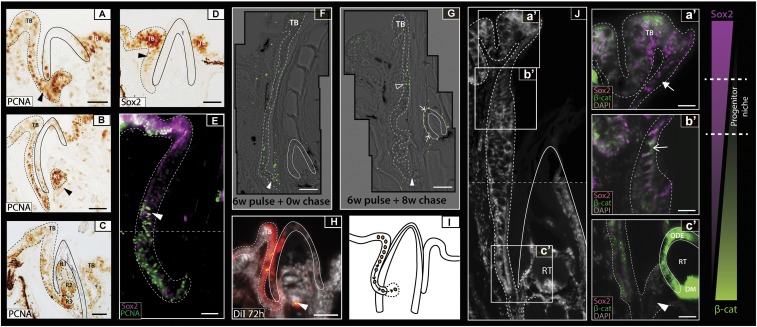
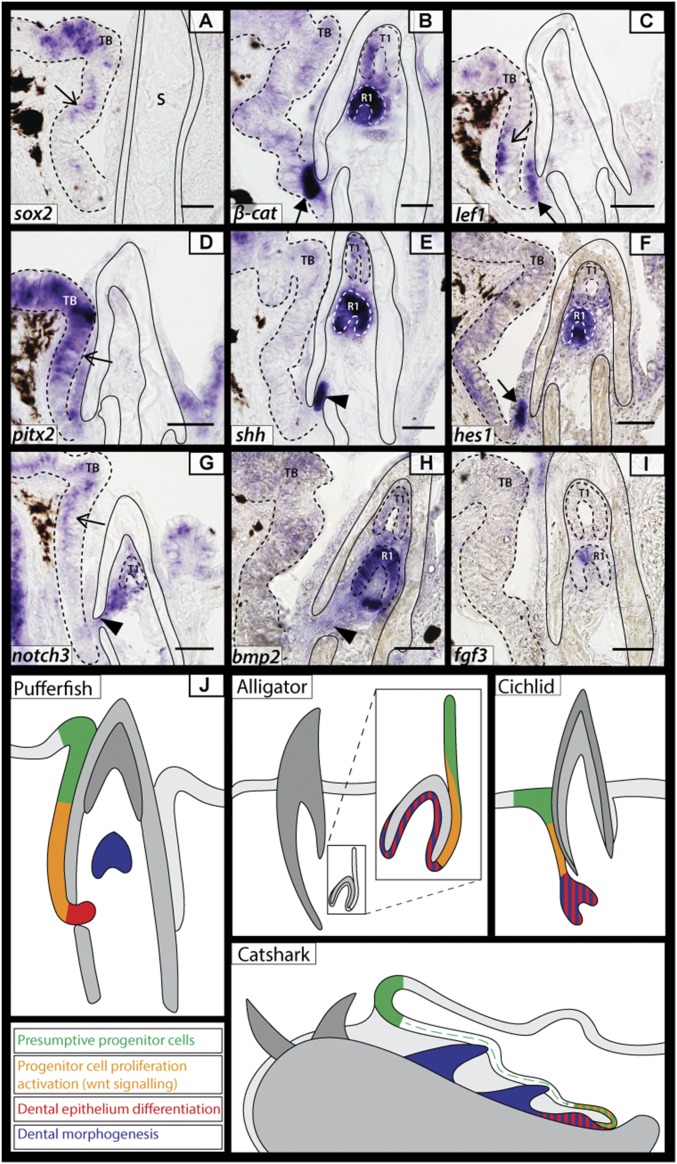
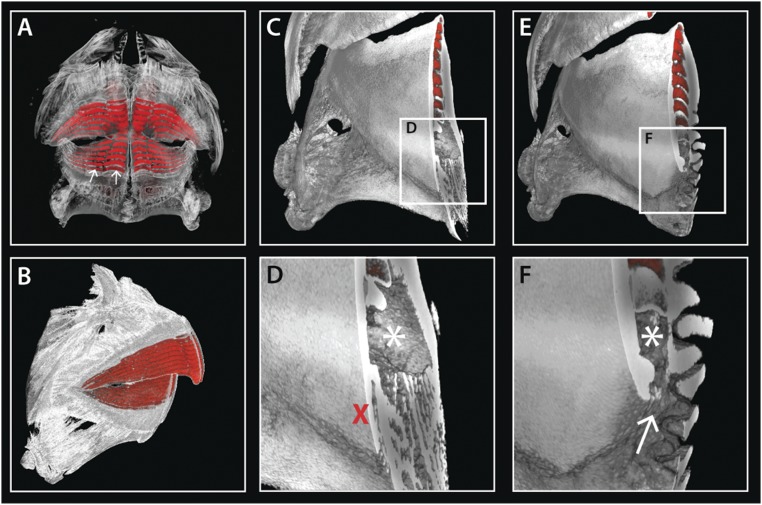
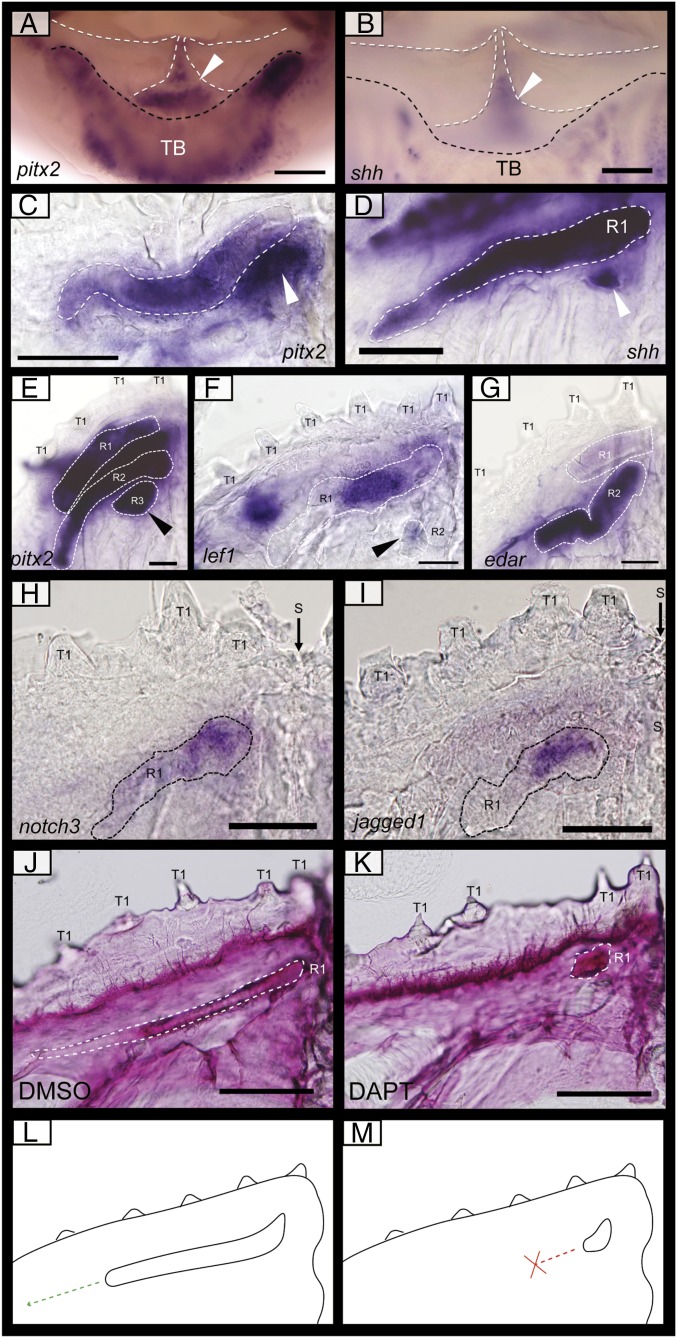
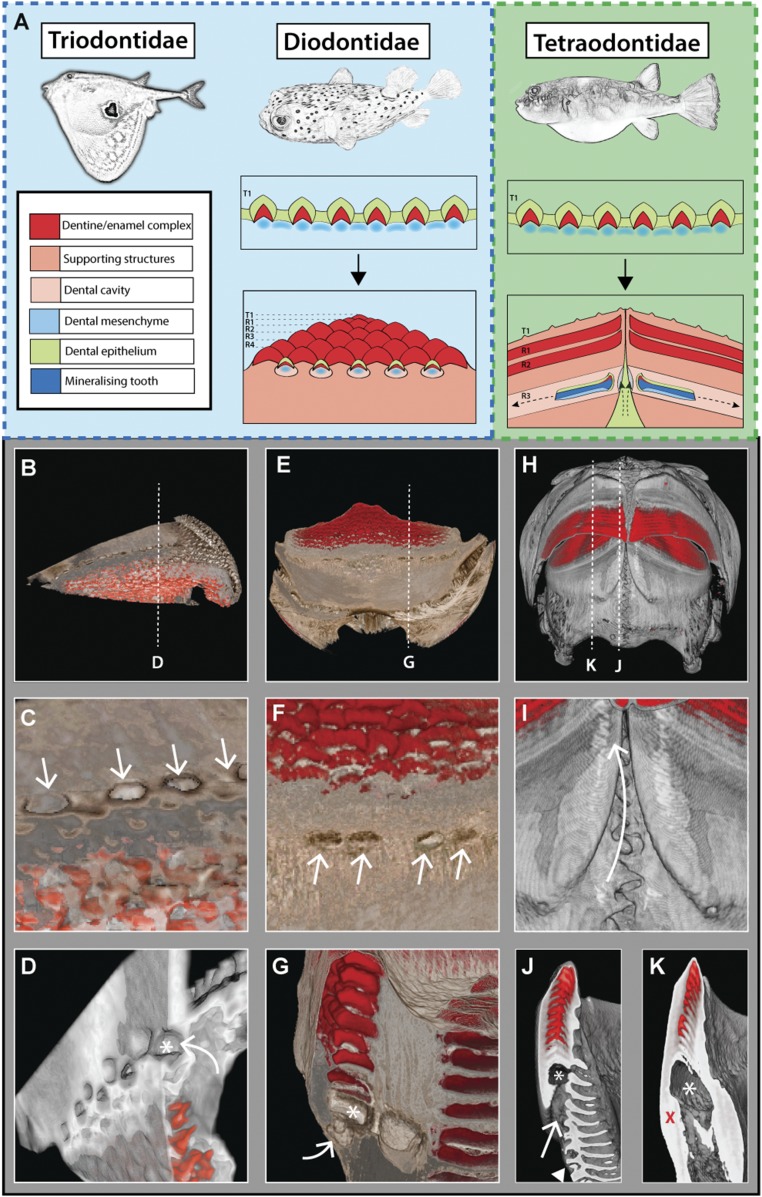
Vertebrate dentitions are extraordinarily diverse in both morphology and regenerative capacity. The teleost order Tetraodontiformes exhibits an exceptional array of novel dental morphologies, epitomized by constrained beak-like dentitions in several families, i.e., porcupinefishes, three-toothed pufferfishes, ocean sunfishes, and pufferfishes. Modification of tooth replacement within these groups leads to the progressive accumulation of tooth generations, underlying the structure of their beaks. We focus on the dentition of the pufferfish (Tetraodontidae) because of its distinct dental morphology. This complex dentition develops as a result of (i) a reduction in the number of tooth positions from seven to one per quadrant during the transition from first to second tooth generations and (ii) a dramatic shift in tooth morphogenesis following the development of the first-generation teeth, leading to the elongation of dental units along the jaw. Gene expression and 1,1'-Dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate (DiI) lineage tracing reveal a putative dental epithelial progenitor niche, suggesting a highly conserved mechanism for tooth regeneration despite the development of a unique dentition. MicroCT analysis reveals restricted labial openings in the beak, through which the dental epithelium (lamina) invades the cavity of the highly mineralized beak. Reduction in the number of replacement tooth positions coincides with the development of only four labial openings in the pufferfish beak, restricting connection of the oral epithelium to the dental cavity. Our data suggest the spatial restriction of dental regeneration, coupled with the unique extension of the replacement dental units throughout the jaw, are primary contributors to the evolution and development of this unique beak-like dentition.
Keywords: dental regeneration; diversity; novelty; stem cells; tooth development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Replacing the first-generation dentition in pufferfish with a unique beak.Proc Natl Acad Sci U S A. 2012 May 22;109(21):8179-84. doi: 10.1073/pnas.1119635109. Epub 2012 May 7. Proc Natl Acad Sci U S A. 2012. PMID: 22566613 Free PMC article.
-
An ancient dental gene set governs development and continuous regeneration of teeth in sharks.Dev Biol. 2016 Jul 15;415(2):347-370. doi: 10.1016/j.ydbio.2016.01.038. Epub 2016 Feb 1. Dev Biol. 2016. PMID: 26845577
-
Common developmental pathways link tooth shape to regeneration.Dev Biol. 2013 May 15;377(2):399-414. doi: 10.1016/j.ydbio.2013.02.007. Epub 2013 Feb 17. Dev Biol. 2013. PMID: 23422830 Free PMC article.
-
A curriculum vitae of teeth: evolution, generation, regeneration.Int J Biol Sci. 2009;5(3):226-43. doi: 10.7150/ijbs.5.226. Epub 2009 Feb 24. Int J Biol Sci. 2009. PMID: 19266065 Free PMC article. Review.
-
The genetic basis of modularity in the development and evolution of the vertebrate dentition.Philos Trans R Soc Lond B Biol Sci. 2001 Oct 29;356(1414):1633-53. doi: 10.1098/rstb.2001.0917. Philos Trans R Soc Lond B Biol Sci. 2001. PMID: 11604128 Free PMC article. Review.
Cited by
-
Re-evaluating the morphological evidence for the re-evolution of lost mandibular teeth in frogs.Evolution. 2021 Dec;75(12):3203-3213. doi: 10.1111/evo.14379. Epub 2021 Nov 9. Evolution. 2021. PMID: 34674263 Free PMC article.
-
Oral and Palatal Dentition of Axolotl Arises From a Common Tooth-Competent Zone Along the Ecto-Endodermal Boundary.Front Cell Dev Biol. 2021 Jan 11;8:622308. doi: 10.3389/fcell.2020.622308. eCollection 2020. Front Cell Dev Biol. 2021. PMID: 33505974 Free PMC article.
-
Count Me in, Count Me out: Regulation of the Tooth Number via Three Directional Developmental Patterns.Int J Mol Sci. 2023 Oct 11;24(20):15061. doi: 10.3390/ijms242015061. Int J Mol Sci. 2023. PMID: 37894742 Free PMC article. Review.
-
Incompatibility between two major innovations shaped the diversification of fish feeding mechanisms.PLoS Biol. 2025 Jun 24;23(6):e3003225. doi: 10.1371/journal.pbio.3003225. eCollection 2025 Jun. PLoS Biol. 2025. PMID: 40554461 Free PMC article.
-
Grand Challenges in Comparative Tooth Biology.Integr Comp Biol. 2020 Sep 1;60(3):563-580. doi: 10.1093/icb/icaa038. Integr Comp Biol. 2020. PMID: 32533826 Free PMC article.
References
-
- Wagner GP, Lynch VJ. Evolutionary novelties. Curr Biol. 2010;20:R48–R52. - PubMed
-
- Jernvall J, Thesleff I. Tooth shape formation and tooth renewal: Evolving with the same signals. Development. 2012;139:3487–3497. - PubMed
-
- Sire JY, Davit-Beal T, Delgado S, Van Der Heyden C, Huysseune A. First-generation teeth in nonmammalian lineages: Evidence for a conserved ancestral character? Microsc Res Tech. 2002;59:408–434. - PubMed
-
- Tucker AS, Fraser GJ. Evolution and developmental diversity of tooth regeneration. Semin Cell Dev Biol. 2014;25-26:71–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
