

Cryo-EM structure of human adenovirus D26 reveals the conservation of structural organization among human adenoviruses
- PMID: 28508067
- PMCID: PMC5425241
- DOI: 10.1126/sciadv.1602670
Cryo-EM structure of human adenovirus D26 reveals the conservation of structural organization among human adenoviruses
Abstract
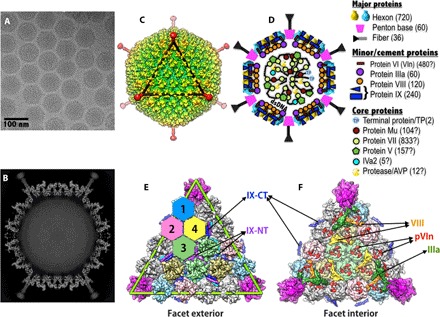
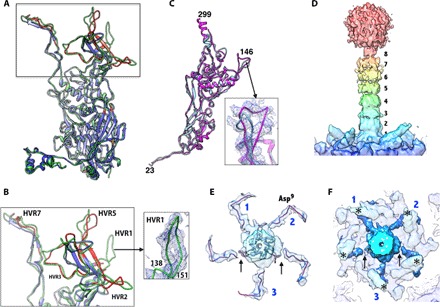
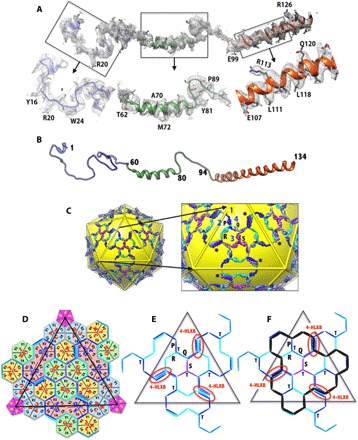
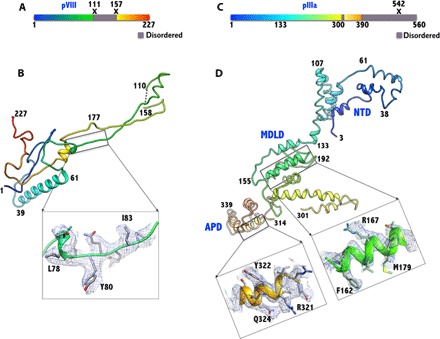
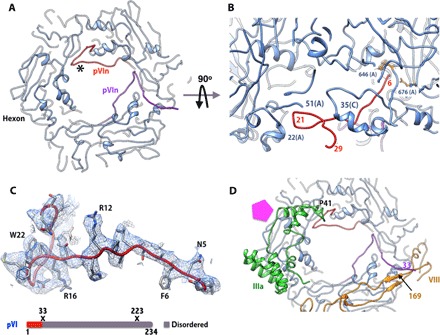
Human adenoviruses (HAdVs) cause acute respiratory, ocular, and gastroenteric diseases and are also frequently used as gene and vaccine delivery vectors. Unlike the archetype human adenovirus C5 (HAdV-C5), human adenovirus D26 (HAdV-D26) belongs to species-D HAdVs, which target different cellular receptors, and is differentially recognized by immune surveillance mechanisms. HAdV-D26 is being championed as a lower seroprevalent vaccine and oncolytic vector in preclinical and human clinical studies. To understand the molecular basis for their distinct biological properties and independently validate the structures of minor proteins, we determined the first structure of species-D HAdV at 3.7 Å resolution by cryo-electron microscopy. All the hexon hypervariable regions (HVRs), including HVR1, have been identified and exhibit a distinct organization compared to those of HAdV-C5. Despite the differences in the arrangement of helices in the coiled-coil structures, protein IX molecules form a continuous hexagonal network on the capsid exterior. In addition to the structurally conserved region (3 to 300) of IIIa, we identified an extra helical domain comprising residues 314 to 390 that further stabilizes the vertex region. Multiple (two to three) copies of the cleaved amino-terminal fragment of protein VI (pVIn) are observed in each hexon cavity, suggesting that there could be ≥480 copies of VI present in HAdV-D26. In addition, a localized asymmetric reconstruction of the vertex region provides new details of the three-pronged "claw hold" of the trimeric fiber and its interactions with the penton base. These observations resolve the previous conflicting assignments of the minor proteins and suggest the likely conservation of their organization across different HAdVs.
Keywords: Ad26; Cement proteins; Cryo-electron microscopy; HAdV-D26; Human adenovirus D26; Major capsid proteins; Minor capsid proteins; Structure.
Figures
References
-
- Davison A. J., Benkő M., Harrach B., Genetic content and evolution of adenoviruses. J. Gen. Virol. 84, 2895–2908 (2003). - PubMed
-
- Rux J. J., Burnett R. M., Type-specific epitope locations revealed by X-ray crystallographic study of adenovirus type 5 hexon. Mol. Ther. 1, 18–30 (2000). - PubMed
-
- Abbink P., Lemckert A. A. C., Ewald B. A., Lynch D. M., Denholtz M., Smits S., Holterman L., Damen I., Vogels R., Thorner A. R., O’Brien K. L., Carville A., Mansfield K. G., Goudsmit J., Havenga M. J. E., Barouch D. H., Comparative seroprevalence and immunogenicity of six rare serotype recombinant adenovirus vaccine vectors from subgroups B and D. J. Virol. 81, 4654–4663 (2007). - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
