

Structures of closed and open conformations of dimeric human ATM
- PMID: 28508083
- PMCID: PMC5425235
- DOI: 10.1126/sciadv.1700933
Structures of closed and open conformations of dimeric human ATM
Abstract
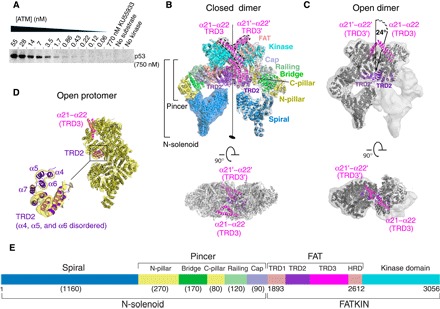
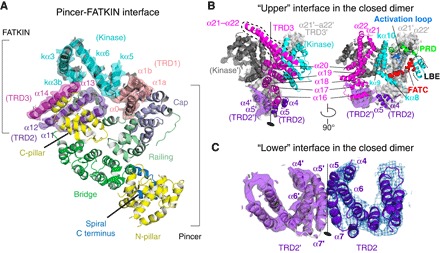
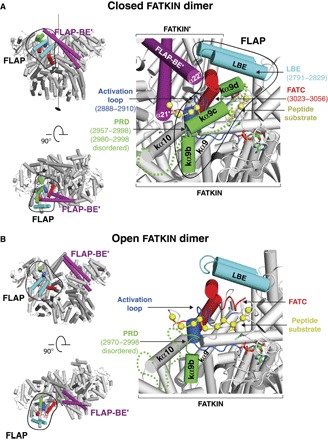
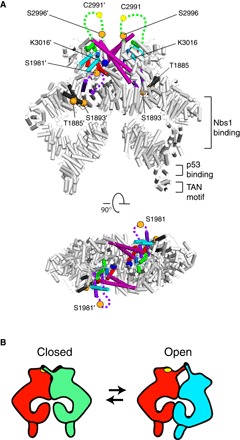
ATM (ataxia-telangiectasia mutated) is a phosphatidylinositol 3-kinase-related protein kinase (PIKK) best known for its role in DNA damage response. ATM also functions in oxidative stress response, insulin signaling, and neurogenesis. Our electron cryomicroscopy (cryo-EM) suggests that human ATM is in a dynamic equilibrium between closed and open dimers. In the closed state, the PIKK regulatory domain blocks the peptide substrate-binding site, suggesting that this conformation may represent an inactive or basally active enzyme. The active site is held in this closed conformation by interaction with a long helical hairpin in the TRD3 (tetratricopeptide repeats domain 3) domain of the symmetry-related molecule. The open dimer has two protomers with only a limited contact interface, and it lacks the intermolecular interactions that block the peptide-binding site in the closed dimer. This suggests that the open conformation may be more active. The ATM structure shows the detailed topology of the regulator-interacting N-terminal helical solenoid. The ATM conformational dynamics shown by the structures represent an important step in understanding the enzyme regulation.
Keywords: ATM; Ataxia telangiectasia mutated; DNA damage response; cryo-EM; kinase; structure.
Figures
References
-
- Shiloh Y., Ziv Y., The ATM protein kinase: Regulating the cellular response to genotoxic stress, and more. Nat. Rev. Mol. Cell. Biol. 14, 197–210 (2013). - PubMed
-
- Bakkenist C. J., Kastan M. B., DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421, 499–506 (2003). - PubMed
-
- Savitsky K., Sfez S., Tagle D. A., Ziv Y., Sartiel A., Collins F. S., Shiloh Y., Rotman G., The complete sequence of the coding region of the ATM gene reveals similarity to cell cycle regulators in different species. Hum. Mol. Genet. 4, 2025–2032 (1995). - PubMed
-
- Paull T. T., Mechanisms of ATM activation. Annu. Rev. Biochem. 84, 711–738 (2015). - PubMed
-
- Mavrou A., Tsangaris G. T., Roma E., Kolialexi A., The ATM gene and ataxia telangiectasia. Anticancer Res. 28, 401–405 (2008). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
