

Efficient Reverse Genetic Systems for Rapid Genetic Manipulation of Emergent and Preemergent Infectious Coronaviruses
- PMID: 28508214
- PMCID: PMC7120940
- DOI: 10.1007/978-1-4939-6964-7_5
Efficient Reverse Genetic Systems for Rapid Genetic Manipulation of Emergent and Preemergent Infectious Coronaviruses
Abstract
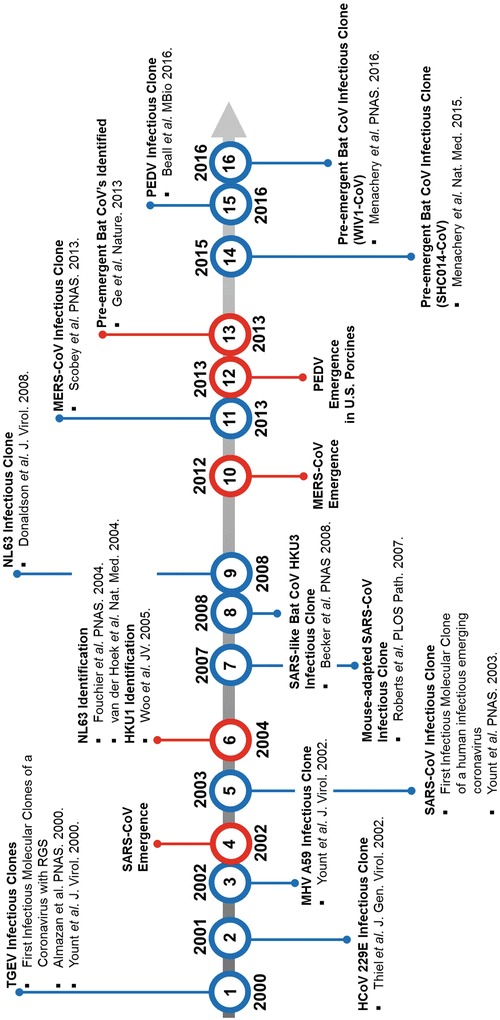
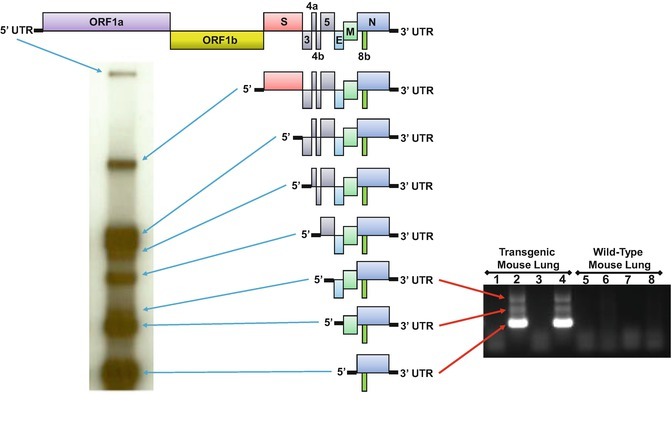
Emergent and preemergent coronaviruses (CoVs) pose a global threat that requires immediate intervention. Rapid intervention necessitates the capacity to generate, grow, and genetically manipulate infectious CoVs in order to rapidly evaluate pathogenic mechanisms, host and tissue permissibility, and candidate antiviral therapeutic efficacy. CoVs encode the largest viral RNA genomes at about 28-32,000 nucleotides in length, and thereby complicate efficient engineering of the genome. Deconstructing the genome into manageable fragments affords the plasticity necessary to rapidly introduce targeted genetic changes in parallel and assort mutated fragments while maximizing genome stability over time. In this protocol we describe a well-developed reverse genetic platform strategy for CoVs that is comprised of partitioning the viral genome into 5-7 independent DNA fragments (depending on the CoV genome), each subcloned into a plasmid for increased stability and ease of genetic manipulation and amplification. Coronavirus genomes are conveniently partitioned by introducing type IIS or IIG restriction enzyme recognition sites that confer directional cloning. Since each restriction site leaves a unique overhang between adjoining fragments, reconstruction of the full-length genome can be achieved through a standard DNA ligation comprised of equal molar ratios of each fragment. Using this method, recombinant CoVs can be rapidly generated and used to investigate host range, gene function, pathogenesis, and candidate therapeutics for emerging and preemergent CoVs both in vitro and in vivo.
Keywords: Bat coronavirus; Coronavirus (CoV); Emerging; Middle East respiratory syndrome coronavirus (MERS-CoV); Porcine epidemic diarrhea virus (PEDV); Preemergent; Reverse genetics; Severe acute respiratory syndrome coronavirus (SARS-CoV).
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
