

Plastic roles of pericytes in the blood-retinal barrier
- PMID: 28508859
- PMCID: PMC5440855
- DOI: 10.1038/ncomms15296
Plastic roles of pericytes in the blood-retinal barrier
Abstract
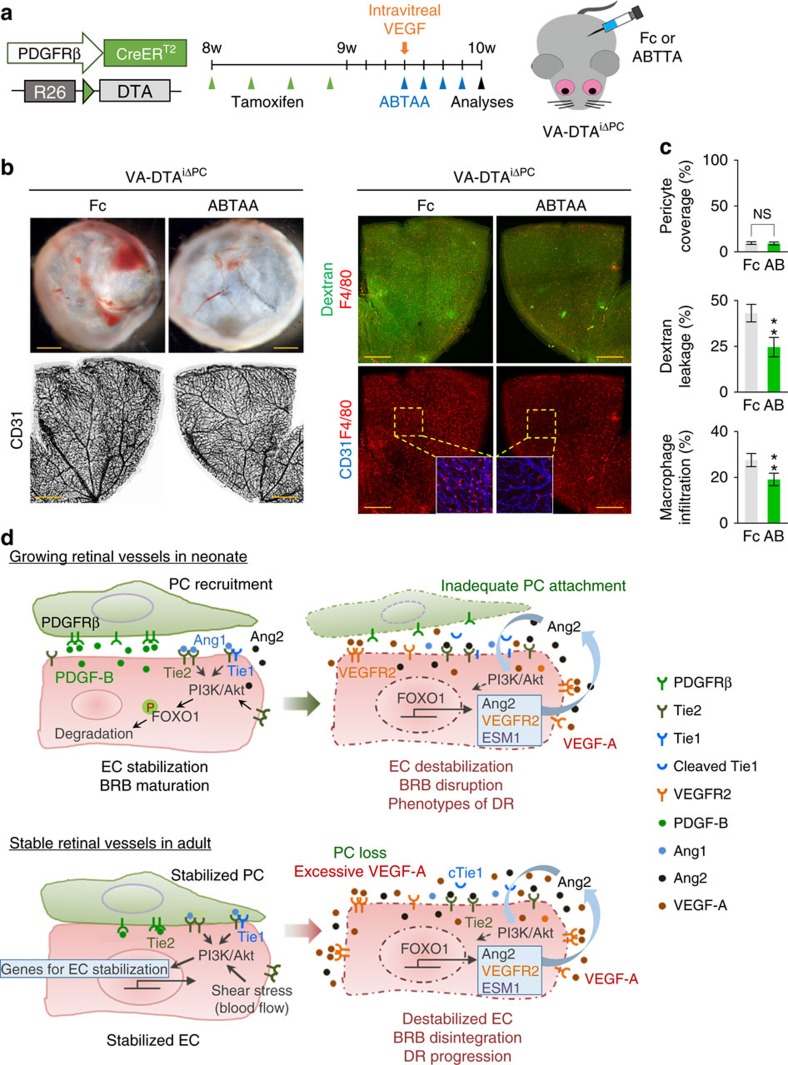
The blood-retinal barrier (BRB) consists of tightly interconnected capillary endothelial cells covered with pericytes and glia, but the role of the pericytes in BRB regulation is not fully understood. Here, we show that platelet-derived growth factor (PDGF)-B/PDGF receptor beta (PDGFRβ) signalling is critical in formation and maturation of BRB through active recruitment of pericytes onto growing retinal vessels. Impaired pericyte recruitment to the vessels shows multiple vascular hallmarks of diabetic retinopathy (DR) due to BRB disruption. However, PDGF-B/PDGFRβ signalling is expendable for maintaining BRB integrity in adult mice. Although selective pericyte loss in stable adult retinal vessels surprisingly does not cause BRB disintegration, it sensitizes retinal vascular endothelial cells (ECs) to VEGF-A, leading to upregulation of angiopoietin-2 (Ang2) in ECs through FOXO1 activation and triggering a positive feedback that resembles the pathogenesis of DR. Accordingly, either blocking Ang2 or activating Tie2 greatly attenuates BRB breakdown, suggesting potential therapeutic approaches to reduce retinal damages upon DR progression.
Conflict of interest statement
The authors declare no competing financial interests.
Figures









Comment in
-
Role of pericytes in the retina.Eye (Lond). 2018 Mar;32(3):483-486. doi: 10.1038/eye.2017.220. Epub 2017 Nov 10. Eye (Lond). 2018. PMID: 29125148 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
