

Thiol redox biochemistry: insights from computer simulations
- PMID: 28509962
- PMCID: PMC5427810
- DOI: 10.1007/s12551-013-0127-x
Thiol redox biochemistry: insights from computer simulations
Abstract
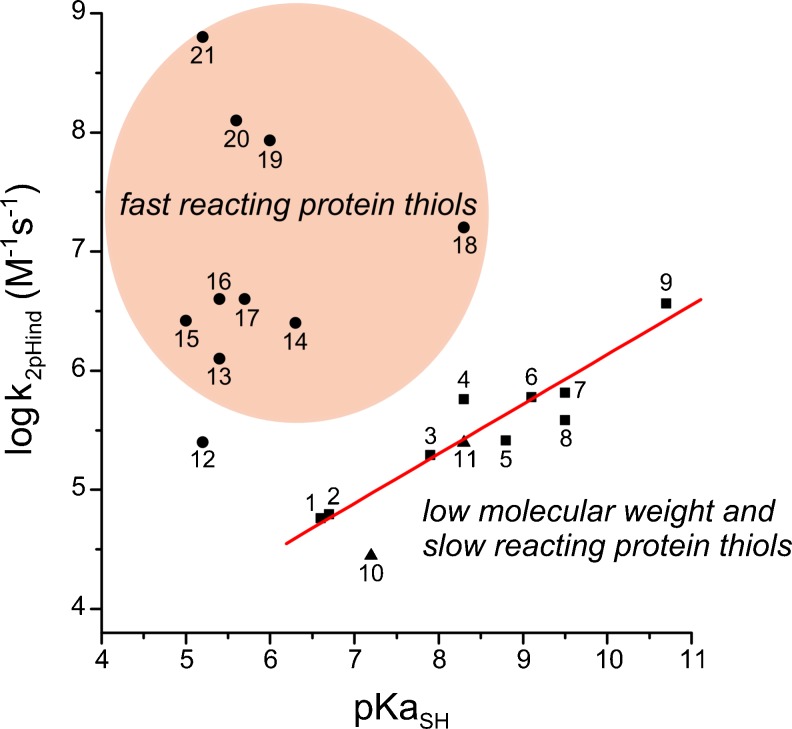
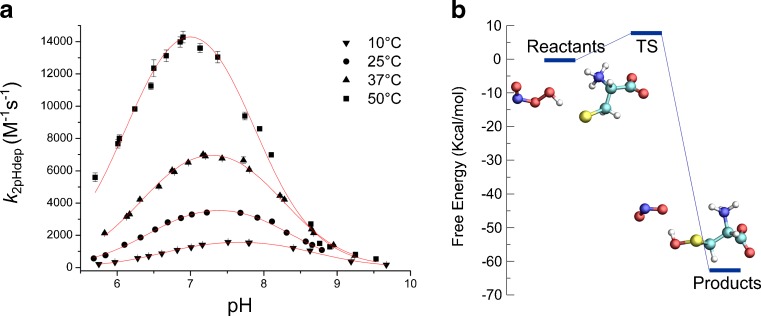
Thiol redox chemical reactions play a key role in a variety of physiological processes, mainly due to the presence of low-molecular-weight thiols and cysteine residues in proteins involved in catalysis and regulation. Specifically, the subtle sensitivity of thiol reactivity to the environment makes the use of simulation techniques extremely valuable for obtaining microscopic insights. In this work we review the application of classical and quantum-mechanical atomistic simulation tools to the investigation of selected relevant issues in thiol redox biochemistry, such as investigations on (1) the protonation state of cysteine in protein, (2) two-electron oxidation of thiols by hydroperoxides, chloramines, and hypochlorous acid, (3) mechanistic and kinetics aspects of the de novo formation of disulfide bonds and thiol-disulfide exchange, (4) formation of sulfenamides, (5) formation of nitrosothiols and transnitrosation reactions, and (6) one-electron oxidation pathways.
Keywords: Computer simulations; Oxidation; Redox homeostasis; Thiols.
Figures



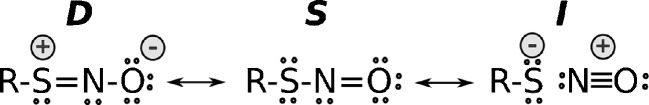


References
-
- Abkevich VI, Shakhnovich EI. What can disulfide bonds tell us about protein energetics, function and folding: simulations and bioninformatics analysis. J Mol Biol. 2000;300:975–985. - PubMed
-
- Ackermann KR, Koster J, Schlücker S. Conformations and vibrational properties of disulfide bridges: Potential energy distribution in the model system diethyl disulfide. Chem Phys. 2009;355:81–84.
-
- Alonso A, Sasin J, Bottini N, Friedberg I, Friedberg I, et al. Protein tyrosine phosphatases in the human genome. Cell. 2004;117:699–711. - PubMed
-
- Anderson BD, Luo D. Application of an exact mathematical model and the steady-state approximation to the kinetics of the reaction of cysteine and hydrogen peroxide in aqueous solution: a reply to the Ashby and Nagy commentary. J Pharm Sci. 2006;95:19–24. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
