

Muscle myosin filaments: cores, crowns and couplings
- PMID: 28509995
- PMCID: PMC5418396
- DOI: 10.1007/s12551-009-0017-4
Muscle myosin filaments: cores, crowns and couplings
Abstract
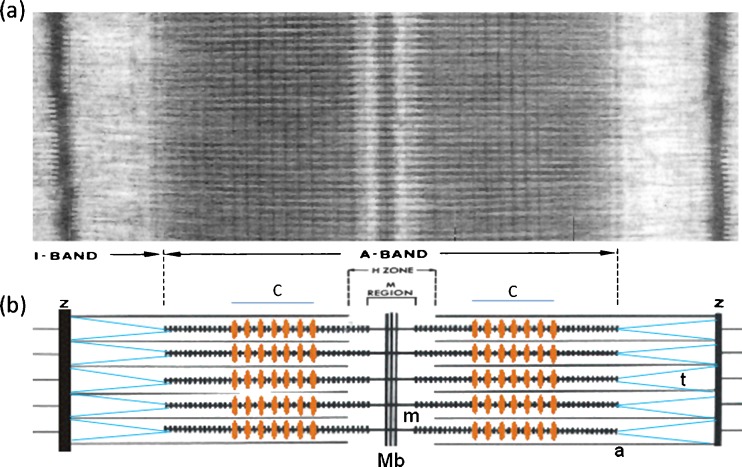
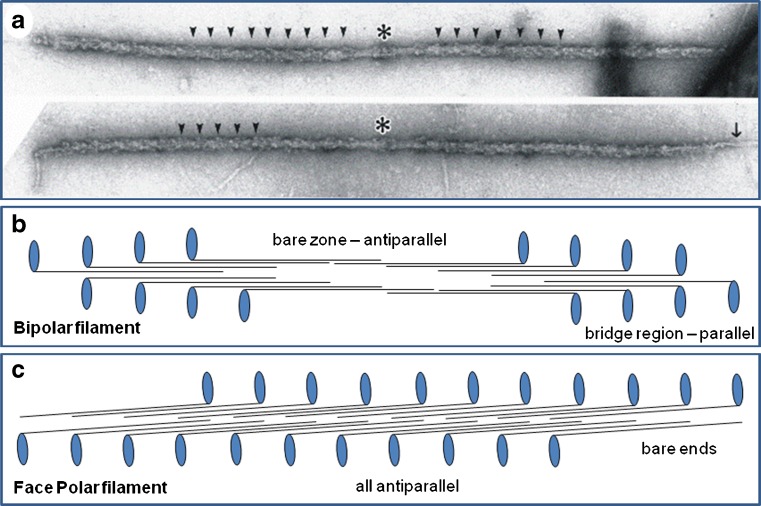
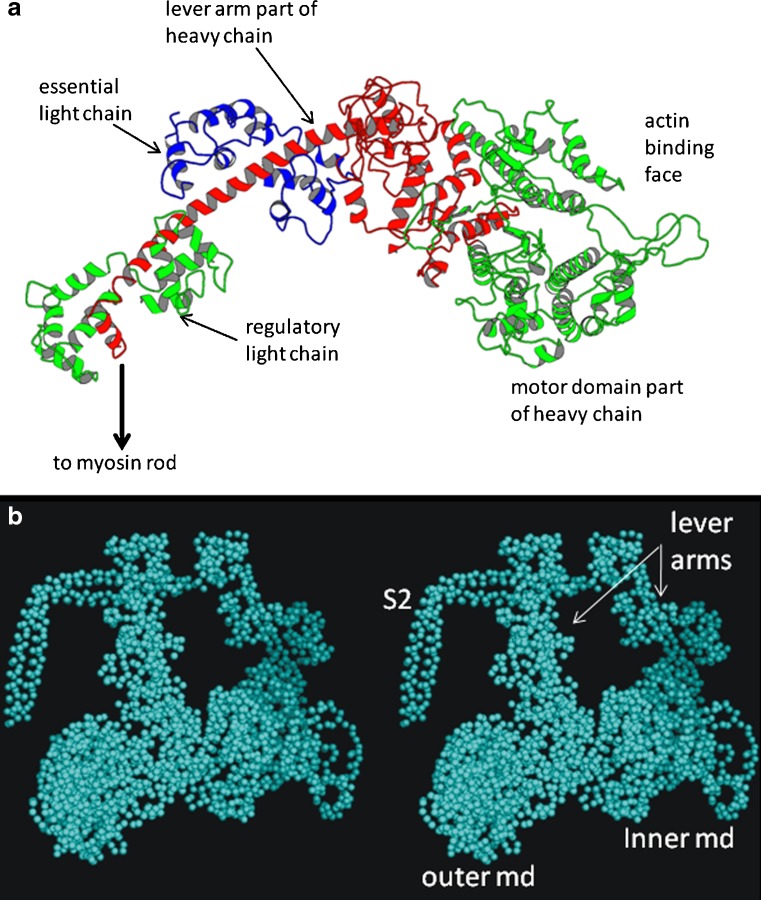
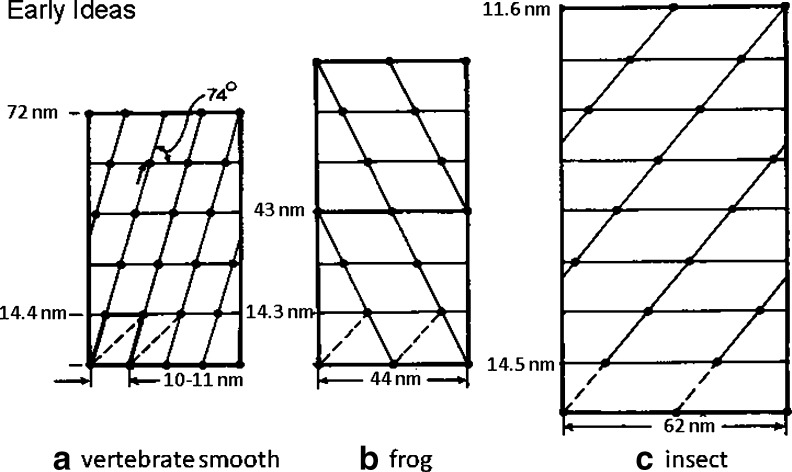
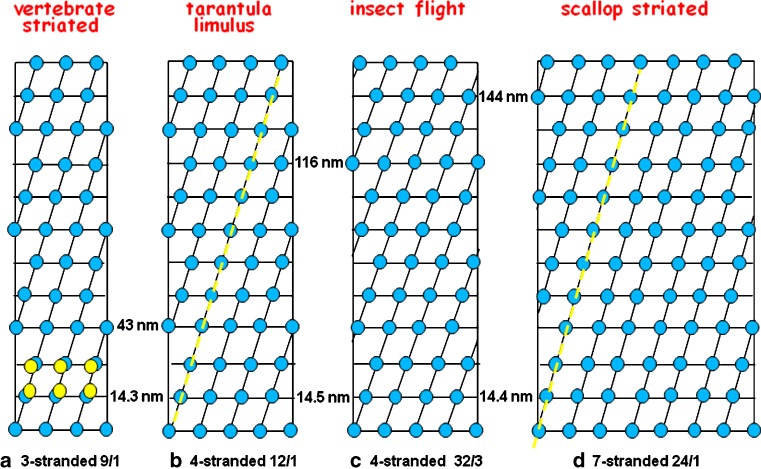
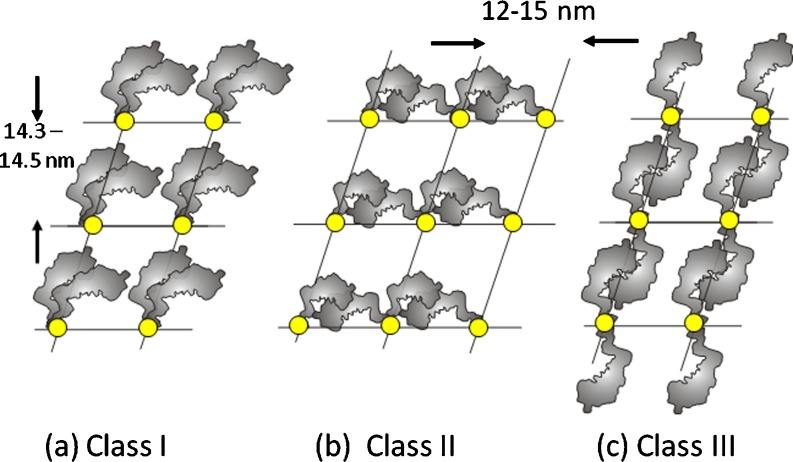
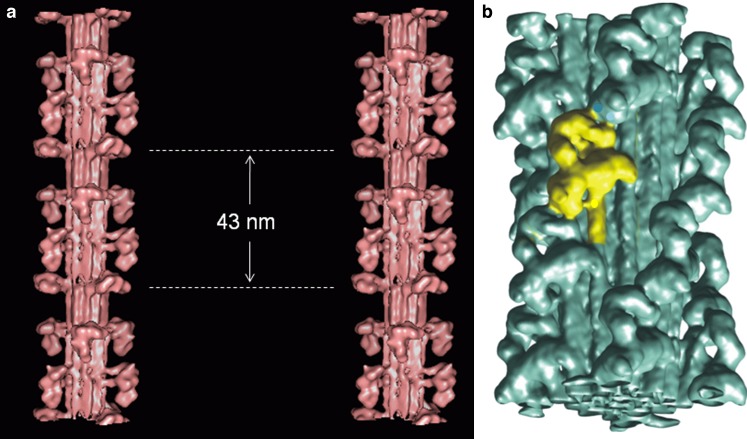
Myosin filaments in muscle, carrying the ATPase myosin heads that interact with actin filaments to produce force and movement, come in multiple varieties depending on species and functional need, but most are based on a common structural theme. The now successful journeys to solve the ultrastructures of many of these myosin filaments, at least at modest resolution, have not been without their false starts and erroneous sidetracks, but the picture now emerging is of both diversity in the rotational symmetries of different filaments and a degree of commonality in the way the myosin heads are organised in resting muscle. Some of the remaining differences may be associated with how the muscle is regulated. Several proteins in cardiac muscle myosin filaments can carry mutations associated with heart disease, so the elucidation of myosin filament structure to understand the effects of these mutations has a clear and topical clinical relevance.
Keywords: Heart disease; Muscle; Myosin filaments; Myosin heads.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
