

The two sides of a lipid-protein story
- PMID: 28510056
- PMCID: PMC5425782
- DOI: 10.1007/s12551-016-0199-5
The two sides of a lipid-protein story
Abstract
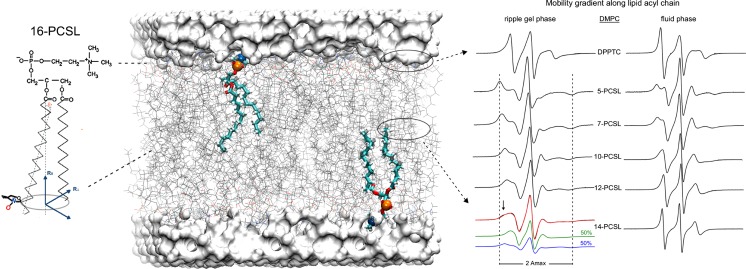
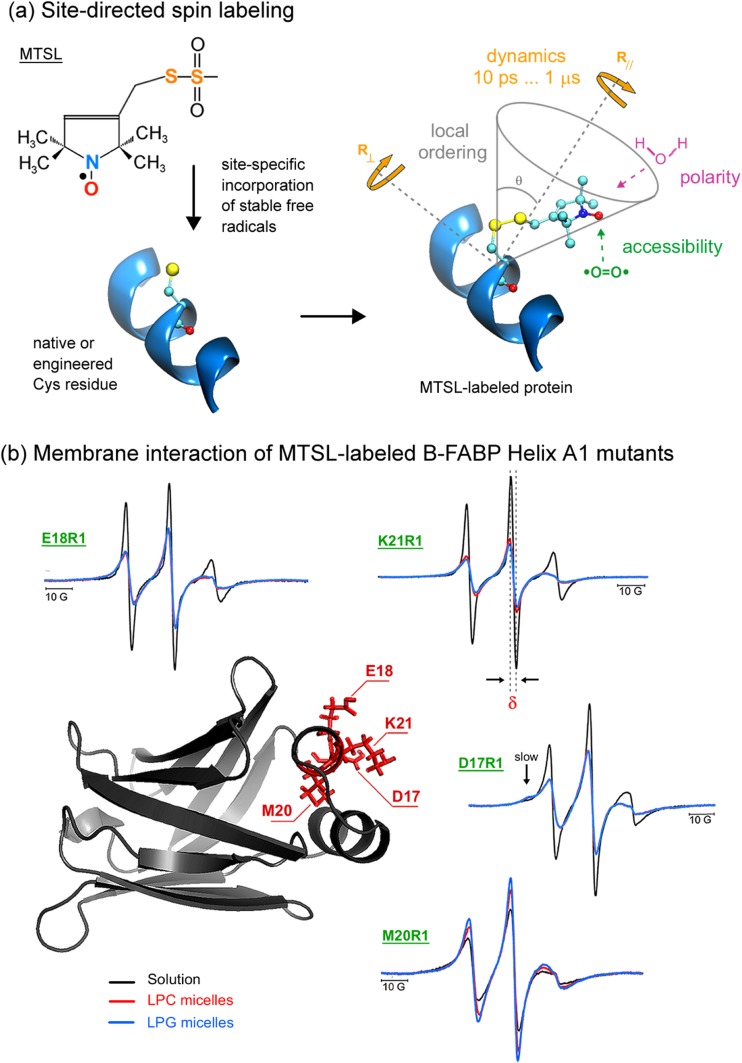
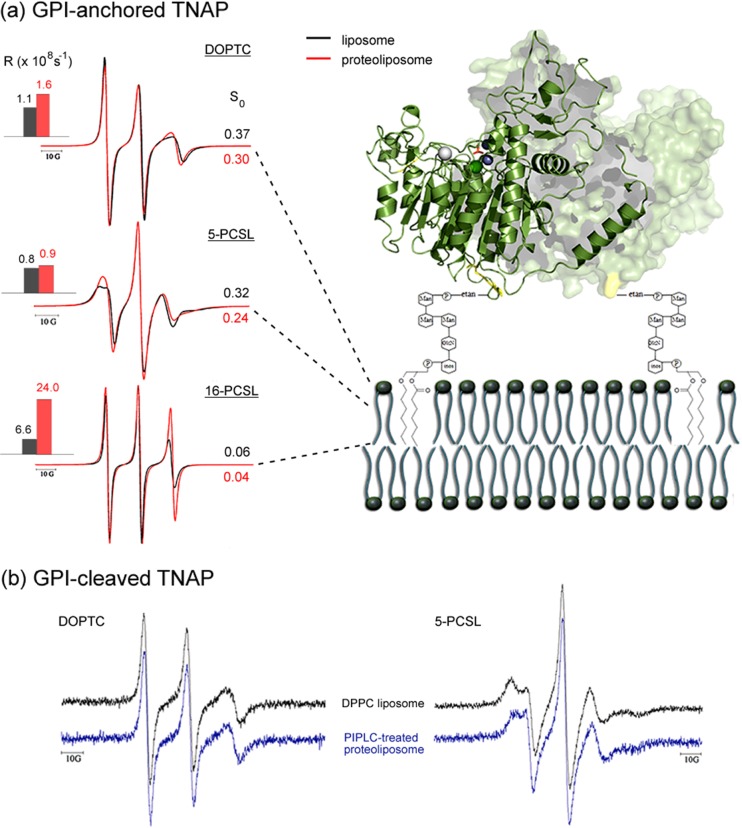
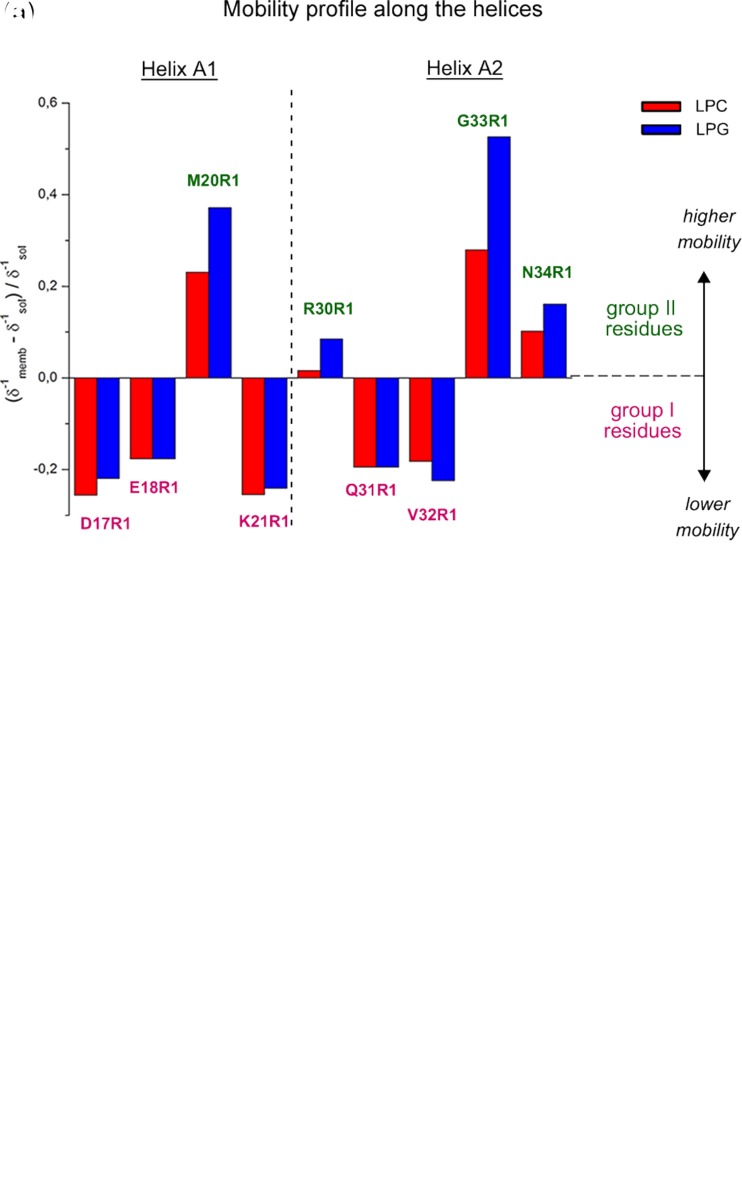
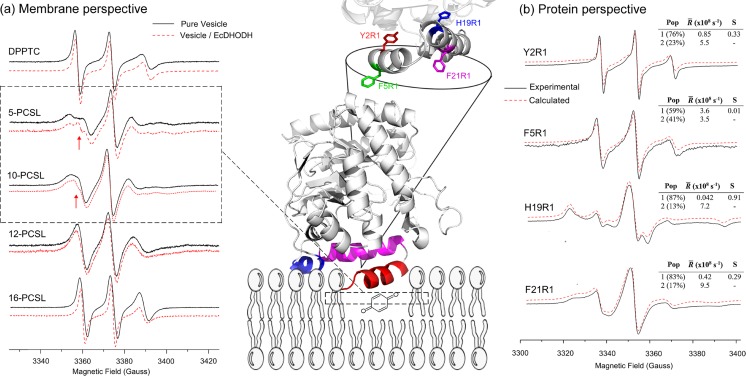
Protein-membrane interactions play essential roles in a variety of cell functions such as signaling, membrane trafficking, and transport. Membrane-recruited cytosolic proteins that interact transiently and interfacially with lipid bilayers perform several of those functions. Experimental techniques capable of probing changes on the structural dynamics of this weak association are surprisingly limited. Among such techniques, electron spin resonance (ESR) has the enormous advantage of providing valuable local information from both membrane and protein perspectives by using intrinsic paramagnetic probes in metalloproteins or by attaching nitroxide spin labels to proteins and lipids. In this review, we discuss the power of ESR to unravel relevant structural and functional details of lipid-peripheral membrane protein interactions with special emphasis on local changes of specific regions of the protein and/or the lipids. First, we show how ESR can be used to investigate the direct interaction between a protein and a particular lipid, illustrating the case of lipid binding into a hydrophobic pocket of chlorocatechol 1,2-dioxygenase, a non-heme iron enzyme responsible for catabolism of aromatic compounds that are industrially released in the environment. In the second case, we show the effects of GPI-anchored tissue-nonspecific alkaline phosphatase, a protein that plays a crucial role in skeletal mineralization, and on the ordering and dynamics of lipid acyl chains. Then, switching to the protein perspective, we analyze the interaction with model membranes of the brain fatty acid binding protein, the major actor in the reversible binding and transport of hydrophobic ligands such as long-chain, saturated, or unsaturated fatty acids. Finally, we conclude by discussing how both lipid and protein views can be associated to address a common question regarding the molecular mechanism by which dihydroorotate dehydrogenase, an essential enzyme for the de novo synthesis of pyrimidine nucleotides, and how it fishes out membrane-embedded quinones to perform its function.
Keywords: EPR; ESR; Metalloproteins; Protein–lipid interaction; Protein–membrane interaction; Spin labeling.
Conflict of interest statement
Conflict of Interest
Luis G. Mansor Basso declares that he has no conflict of interest.
Luis F. Santos Mendes declares that he has no conflict of interest.
Antonio J. Costa-Filho declares that he has no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Figures
References
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K and Walter P (2007) Molecular biology of the cell (5th edn). Garland, New York
-
- Anderson HC. Molecular biology of matrix vesicles. Clin Orthop Relat Res. 1995;314:266–280. - PubMed
-
- Atlas RM, Cerniglia CE. Bioremediation of petroleum pollutants — diversity and environmental aspects of hydrocarbon biodegradation. Bioscience. 1995;45:332–338. doi: 10.2307/1312494. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
