

Measuring the flow of molecules in cells
- PMID: 28510061
- PMCID: PMC5418376
- DOI: 10.1007/s12551-011-0051-x
Measuring the flow of molecules in cells
Abstract
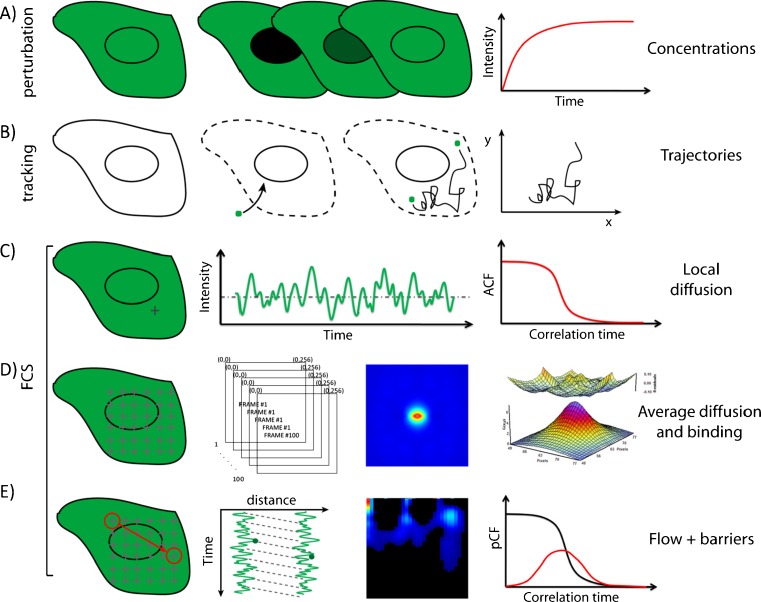
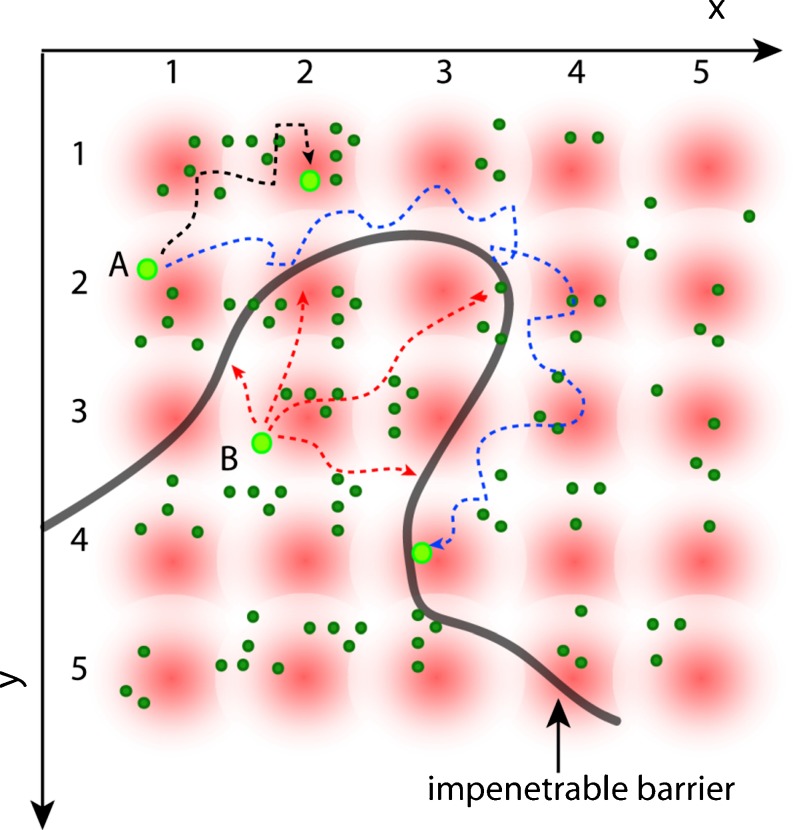
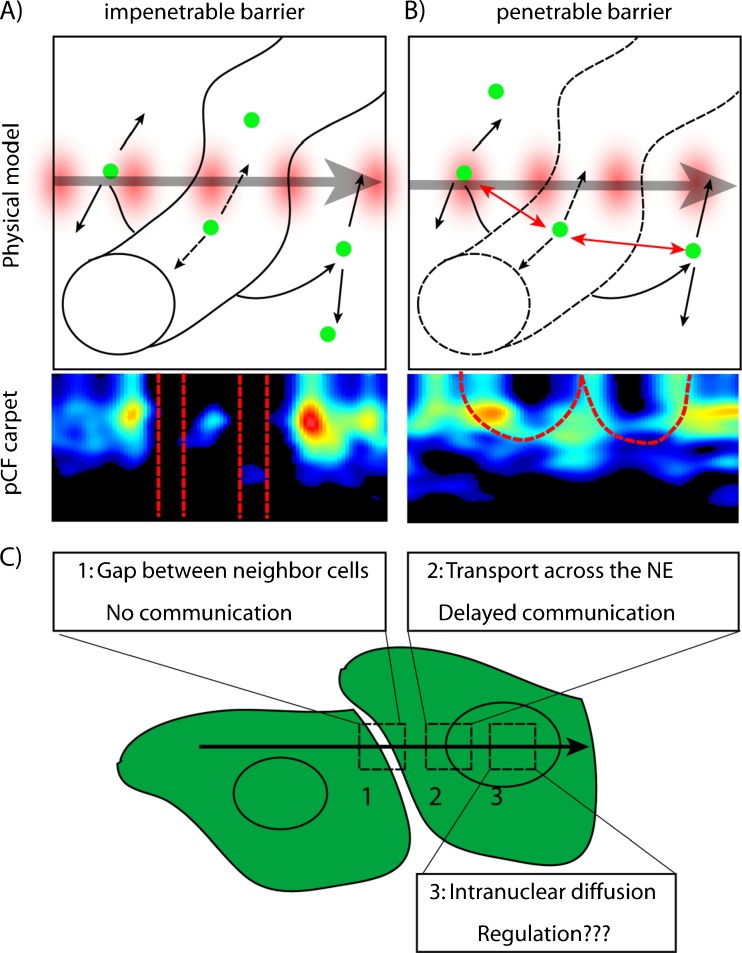
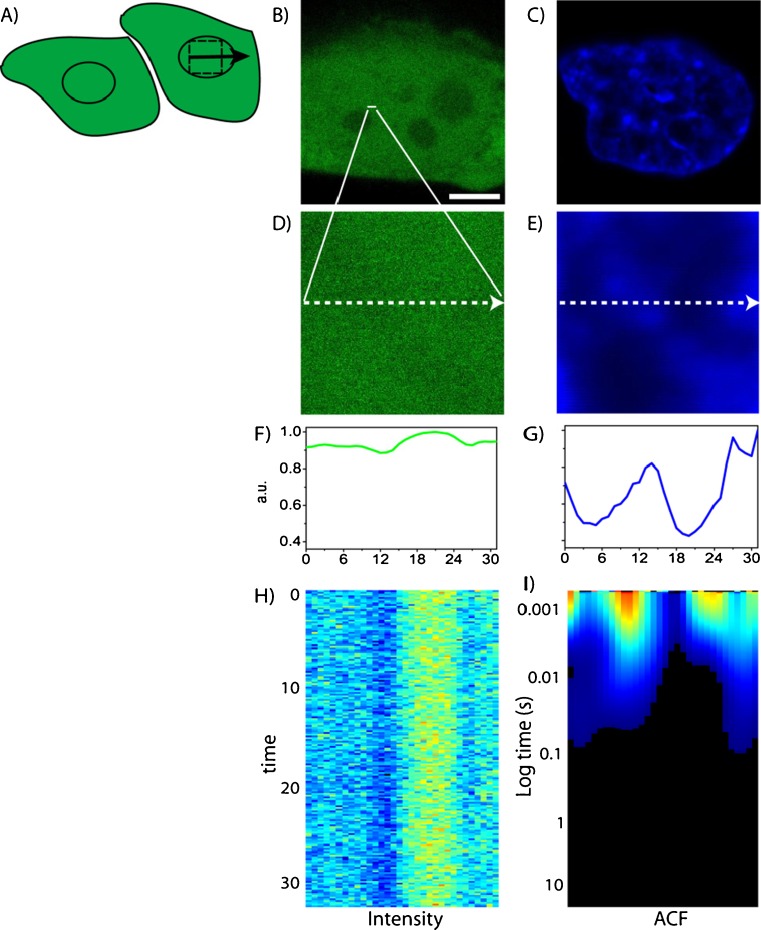
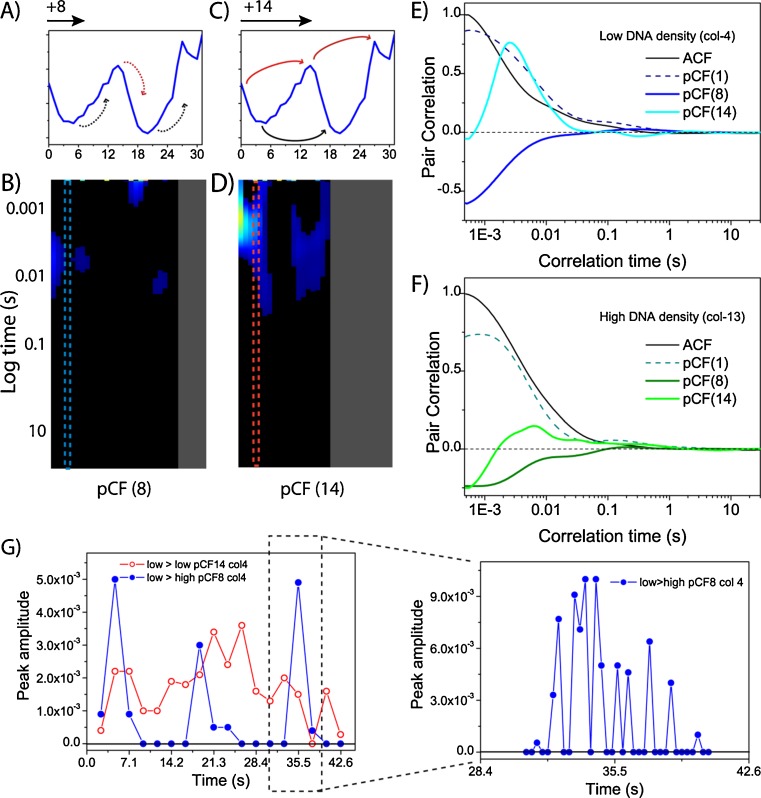
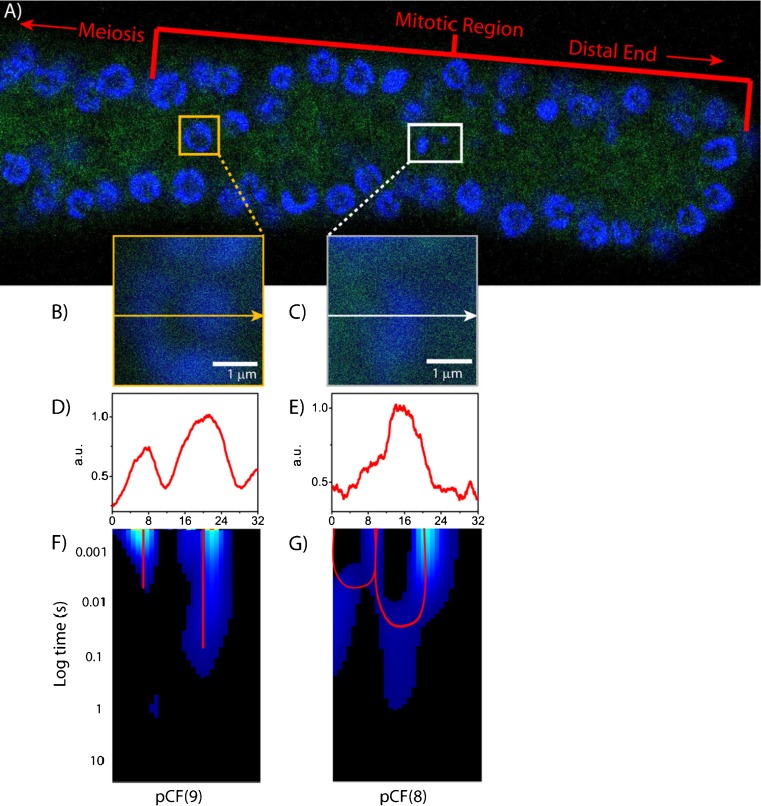
No methods proposed thus far have the capability to measure molecular flow in live cells at the single molecule level. Here, we review the potentiality of a newly established method based on the spatial correlation of fluorescence fluctuations at a pair of points in the sample (pair correlation method). The pair correlation function (pCF) offers a unique tool to probe the directionality of intracellular traffic, by measuring the accessibility of the cellular landscape and its role in determining the diffusive routes adopted by molecules. The sensitivity of the pCF method toward detection of barriers means that different structural elements of the cell can be tested in terms of penetrability and mechanisms of regulation imparted on molecular flow. This has been recently demonstrated in a series of studies looking at molecular transport inside live cells. Here, we will review the theory behind detection of barriers to molecular flow, the rules to interpret pCF data, and relevant applications to intracellular transport.
Keywords: Intracellular transport; Molecular flow; Pair correlation function; Spatial correlation.
Figures

Similar articles
-
Diffusion Tensor Analysis by Two-Dimensional Pair Correlation of Fluorescence Fluctuations in Cells.Biophys J. 2016 Aug 23;111(4):841-851. doi: 10.1016/j.bpj.2016.07.005. Biophys J. 2016. PMID: 27558727 Free PMC article.
-
Visualization of barriers and obstacles to molecular diffusion in live cells by spatial pair-cross-correlation in two dimensions.Biomed Opt Express. 2017 Dec 20;9(1):303-321. doi: 10.1364/BOE.9.000303. eCollection 2018 Jan 1. Biomed Opt Express. 2017. PMID: 29359105 Free PMC article.
-
Imaging barriers to diffusion by pair correlation functions.Biophys J. 2009 Jul 22;97(2):665-73. doi: 10.1016/j.bpj.2009.04.048. Biophys J. 2009. PMID: 19619481 Free PMC article.
-
Flow cytometry: basic principles and applications.Crit Rev Biotechnol. 2017 Mar;37(2):163-176. doi: 10.3109/07388551.2015.1128876. Epub 2016 Jan 14. Crit Rev Biotechnol. 2017. PMID: 26767547 Review.
-
Recent applications of fluorescence correlation spectroscopy in live systems.FEBS Lett. 2014 Oct 1;588(19):3571-84. doi: 10.1016/j.febslet.2014.03.056. Epub 2014 Apr 12. FEBS Lett. 2014. PMID: 24726724 Review.
Cited by
-
Back to the Future: Genetically Encoded Fluorescent Proteins as Inert Tracers of the Intracellular Environment.Int J Mol Sci. 2020 Jun 11;21(11):4164. doi: 10.3390/ijms21114164. Int J Mol Sci. 2020. PMID: 32545175 Free PMC article. Review.
-
Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset.Sci Rep. 2013;3:1141. doi: 10.1038/srep01141. Epub 2013 Jan 25. Sci Rep. 2013. PMID: 23355954 Free PMC article.
-
Changes in chromatin compaction during the cell cycle revealed by micrometer-scale measurement of molecular flow in the nucleus.Biophys J. 2012 Feb 8;102(3):691-7. doi: 10.1016/j.bpj.2011.11.4026. Epub 2012 Feb 7. Biophys J. 2012. PMID: 22325293 Free PMC article.
-
Spatiotemporal regulation of Heterochromatin Protein 1-alpha oligomerization and dynamics in live cells.Sci Rep. 2015 Aug 4;5:12001. doi: 10.1038/srep12001. Sci Rep. 2015. PMID: 26238434 Free PMC article.
-
Stability of cytoplasmic nanoviscosity during cell cycle of HeLa cells synchronized with Aphidicolin.Sci Rep. 2019 Nov 11;9(1):16486. doi: 10.1038/s41598-019-52758-6. Sci Rep. 2019. PMID: 31712575 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
