

Historical perspective on heart function: the Frank-Starling Law
- PMID: 28510104
- PMCID: PMC5418489
- DOI: 10.1007/s12551-015-0184-4
Historical perspective on heart function: the Frank-Starling Law
Abstract
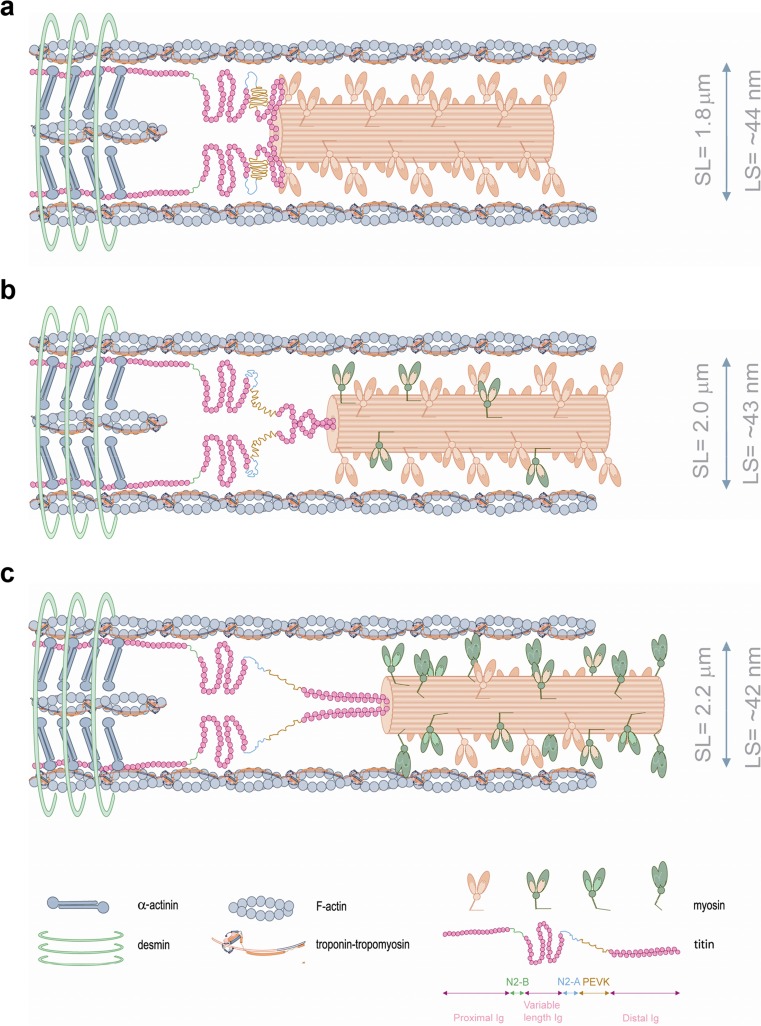
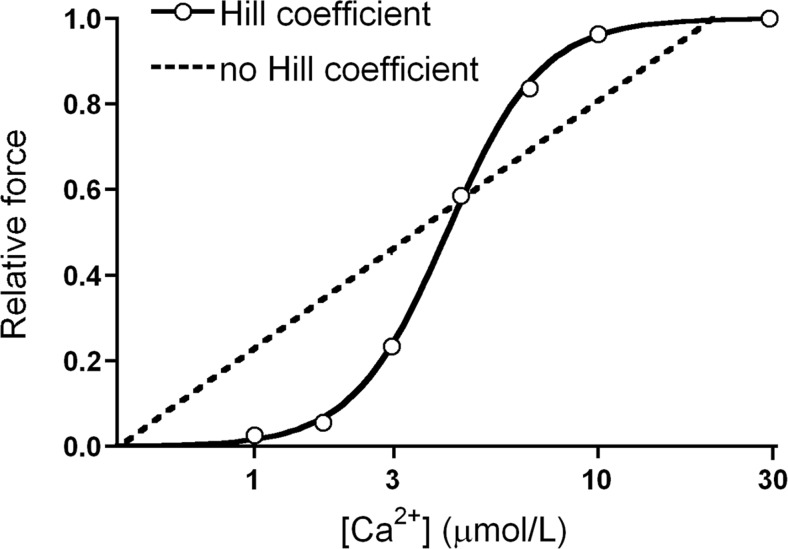
More than a century of research on the Frank-Starling Law has significantly advanced our knowledge about the working heart. The Frank-Starling Law mandates that the heart is able to match cardiac ejection to the dynamic changes occurring in ventricular filling and thereby regulates ventricular contraction and ejection. Significant efforts have been attempted to identify a common fundamental basis for the Frank-Starling heart and, although a unifying idea has still to come forth, there is mounting evidence of a direct relationship between length changes in individual constituents (cardiomyocytes) and their sensitivity to Ca2+ ions. As the Frank-Starling Law is a vital event for the healthy heart, it is of utmost importance to understand its mechanical basis in order to optimize and organize therapeutic strategies to rescue the failing human heart. The present review is a historic perspective on cardiac muscle function. We "revive" a century of scientific research on the heart's fundamental protein constituents (contractile proteins), to their assemblies in the muscle (the sarcomeres), culminating in a thorough overview of the several synergistically events that compose the Frank-Starling mechanism. It is the authors' personal beliefs that much can be gained by understanding the Frank-Starling relationship at the cellular and whole organ level, so that we can finally, in this century, tackle the pathophysiologic mechanisms underlying heart failure.
Keywords: Cardiomyocytes; Frank–Starling; Heart; History; Myofilaments.
Conflict of interest statement
Conflict of interest
Vasco Sequeira declares that he has no conflict of interest.
Jolanda van der Velden declares that she has no conflict of interest.
Ethical approval
This article does not contain any studies with human or animal subjects performed by the authors.
Figures
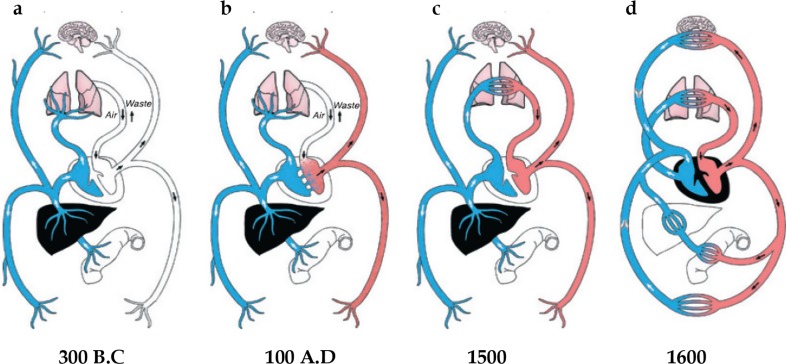
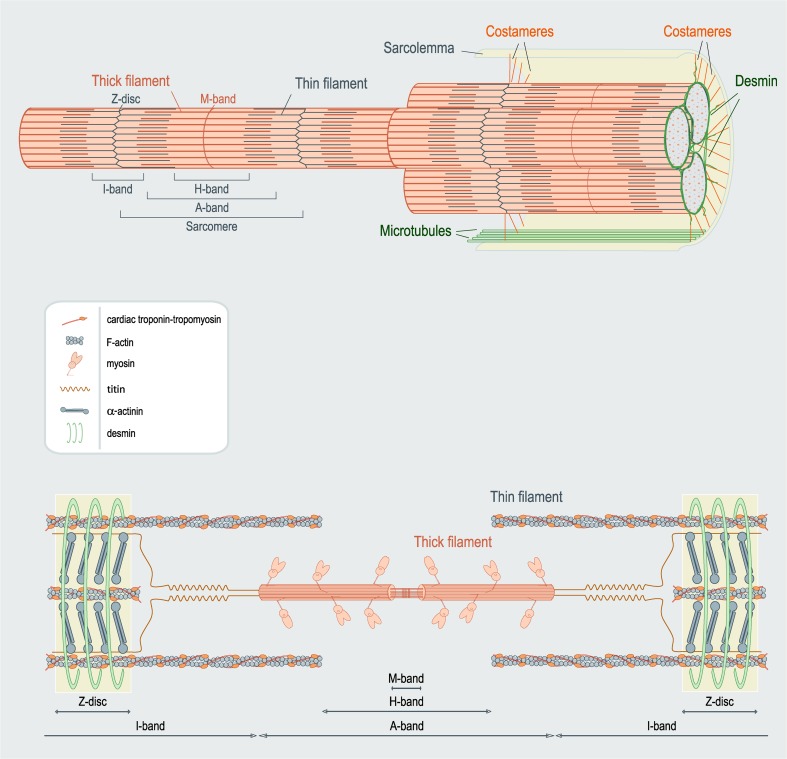
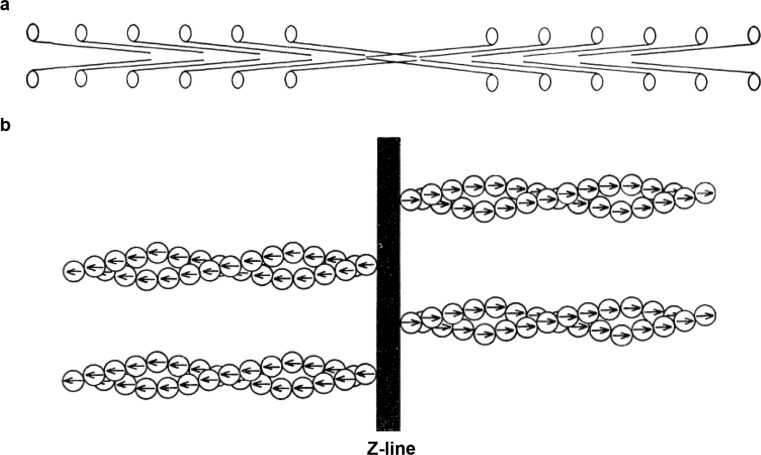
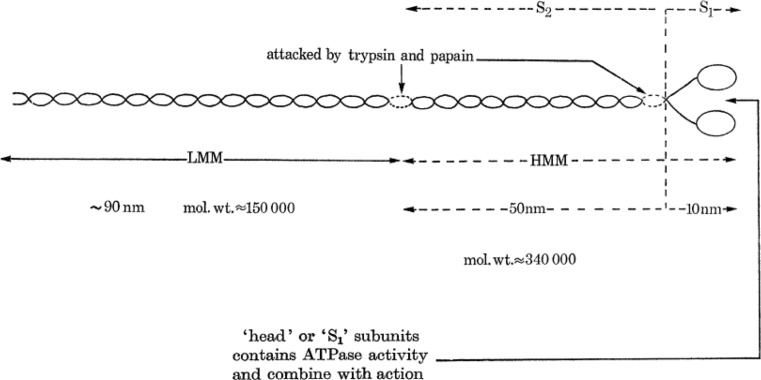




References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
